
Une autre histoire de l'humanité
3. L'évolution de La vie
et les champs d'information
Partie 2
Le darwinisme en question à la lumière de la biologie
Alain Boudet
Dr en Sciences Physiques
Résumé: 2. De plus en plus de chercheurs soulignent les inconsistances de la théorie néodarwiniste, construite sur une interprétation controversée des découvertes archéologiques. L'un des points les plus contestables est que des mutations aléatoires puissent conférer des caractères avantageux. Un autre est que les caractères complexes devraient apparaitre progressivement. Or les bactéries incluent des systèmes biologiques très élaborés qui sont apparus tout faits. Les nombreuses avancées de la biologie montrent que la transmission génétique est bien plus complexe que par la voie unique des gènes. Les zones dites non-codantes jouent un rôle très important dans la régulation génétique et des instructions nouvelles peuvent passer dans la descendance sans qu'il y ait de mutations dans les gènes. Enfin, la composition de l'ADN des mitochondries montre que les espèces sont génétiquement bien distinctes, sans chainon intermédiaire. La synthèse évolutive étendue tient compte de cette complexité et de cette mobilité en intégrant l'épigénétique et la théorie des systèmes.
| Partie 1: Darwin et la théorie moderne de l'évolution
| |
| Contenu de la partie 2
| Annexe |
| Partie 3: Un principe organisateur - Pratiques inquiétantes dans la science
| |
| Partie 4: Information et mythes de création
| |
Dès son apparition, la théorie darwiniste a suscité de nombreuses controverses. Elles se sont poursuivies, ont été renouvelées et reprécisées avec la théorie néodarwiniste (exposée dans la partie précédente). Cette deuxième partie traite des nombreux points de cette théorie qui sont obscurs et contestables.
La théorie néodarwiniste, ou théorie moderne/synthétique de l'évolution, repose sur un ensemble de principes qui s'imbriquent, se complètent et sont tous nécessaires:
Si l'un de ces principes est faux, cela remet en cause tout le système.
Nous allons examiner la fragilité et les failles de ces principes et rapporter quelques pistes alternatives empruntées par d'autres scientifiques.
Selon la théorie néodarwiniste, les mutations génétiques se produisent de façon aléatoire selon une fréquence régulière au cours du temps. Cela implique que les espèces évoluent de façon continue et progressive (théorie dite uniformitariste ou gradualiste). On devrait donc logiquement voir apparaitre de nouvelles espèces à un rythme continu et constant.
Or plus les découvertes paléontologiques se font jour, plus ce schéma uniformitariste est remis en cause.
La réalité fossile visible est que des espèces restent inchangées pendant de longues périodes. Puis des extinctions subites (à l'échelle géologique) se produisent, au cours desquelles de nombreuses espèces disparaissent, parfois de façon massive. Elles sont suivies de brusques explosions de formes de vie. (voir article précédent L'histoire des animaux)

Stephen Gould
Merci à Stephen Jay Gould Archives
Cette réalité archéologique a conduit certains paléontologues à se détacher du schéma uniformitariste et à proposer d'autres scénarios. C'est le cas de deux paléontologues états-uniens, Stephen Jay Gould (1941 - 2002) et Niles Eldredge, qui en 1972 ont proposé la théorie des équilibres ponctués (Punctuated equilibrium: an alternative to phyletic gradualism, dans l'ouvrage collectif Models in Paleobiology, p.82, éd. Th. Schopf).
Cette théorie énonce que les changements évolutifs arrivent par saccades brusques, correspondant aux grandes extinctions. Des apparitions rapides de nouvelles espèces ponctuent de longues périodes d'équilibre (stases).
Des changements continus peuvent en même temps se produire au sein de groupes limités dans l'espace et dans le temps.
Pour Gould, l’histoire de la vie se caractérise par la diversité, de la bactérie aux primates, plus que par un accroissement de la complexité. Nous vivons encore à l'ère des bactéries, qui n'ont pas beaucoup changé depuis leur apparition il y a environ 3,5 milliards d'années. Par leurs capacités extraordinaires, elles sont les reines de la vie. (Entretien avec S. Gould, 01/10/1997, Révolution lente)
La communauté scientifique rejeta majoritairement la théorie des changements brusques, pour des raisons émotionnelles. Elle lui rappelait le catastrophisme promu par les créationnistes en référence aux récits de la Bible, comme le déluge, et il fallait à tout prix éviter toute réminiscence biblique.
Pourtant, la théorie des équilibres ponctuées n'apporte qu'une nouveauté modérée. Déjà suggérée par Darwin lui-même, elle se contente de décrire les observations fossiles en les situant dans le temps - présence des espèces selon des rythmes. Elle ne propose pas d'explication sur le moteur de ces explosions de vie.
La résistance à cette théorie somme toutes peu révolutionnaire révèle combien il est difficile d'apporter des propositions radicalement nouvelles dans ce milieu scientifique. Nous y reviendrons plus en détail dans la partie 3.
Sources: Équilibre ponctué dans Wikipédia
Selon la théorie de l'évolution, la sélection des individus les plus adaptés est un mécanisme fondamental majeur. Il énonce que les espèces qui possèdent les variations génétiques prétendument avantageuses peuvent se développer au détriment des autres, qui disparaissent dans leur lutte pour la survie sur un même territoire.
Cette sélection naturelle est-elle véritablement observée dans la nature? Les auteurs darwinistes affirment que oui et pour l'illustrer, ils citent presque toujours deux cas:
J'ai présenté dans la partie 1 les études faites par Darwin lui-même sur les becs des pinsons des Galapagos. J'ai aussi rapporté une étude récente d'une équipe de scientifiques de Bristol qui a montré que les becs de ces pinsons servent à de multiples fonctions, et qu'il est donc impossible d'imaginer qu'une évolution ait eu lieu sur la base d'un avantage pour une seule de ces fonctions.
Le papillon nocturne nommé phalène du bouleau a été étudié en 1955 par le zoologiste britannique Bernard Kettlewell. On le trouve parfois posé sur le tronc des bouleaux avec lequel il se confond à cause de sa couleur claire (forme de mimétisme).
Or il en existe une seconde espèce aux ailes sombres, remarquée depuis le 19e siècle à proximité des centres industriels anglais. Selon Kettlewell, la cause en est le noircissement des arbres par les fumées industrielles. Les phalènes blanches sont bien plus visibles sur un fond sombre et plus vulnérables vis-à-vis des oiseaux prédateurs. En devenant sombres, elles se parent d'un nouveau camouflage. L'apparition d'une espèce sombre serait donc l'exemple typique d'une mutation rapide due à la pression de l'environnement et à la sélection naturelle.
 |  |  |
| Phalène du bouleau, forme claire, ailes déployées Merci à Chiswick Chap, Wikimedia | Phalène du bouleau, forme sombre, ailes déployées Merci à Chiswick Chap, Wikimedia | Phalène claire en position de repos. Merci à Christian Le Guillou, Académie Aix-Marseille |
En réalité, cette étude souffre de lacunes importantes. La principale est que lorsque les phalènes se posent sur l'arbre, leurs ailes ne sont pas déployées comme le présentent tous les articles, mais repliées, donc avec une teinte différente de celle du bouleau.
Voici ce que dit à ce sujet Christian Le Guillou, inspecteur pédagogique de SVT, académie Aix-Marseille: On s’étonne que [la communauté scientifique] ait si bien accepté des photographies montrant des phalènes en position de repos, ailes déployées, alors que tous les ornithologues s’accordaient déjà à proclamer qu’elles se reposent ailes repliées, privilégiant ainsi comme seul facteur déterminant un changement dans la composition de la population, la prédation exercée par les oiseaux. (La Phalène du bouleau, du fait construit au fait mythifié, Christian Le Guillou, Avril 2009)
Dans une autre critique, un biologiste avance que l'endroit de repos habituel des phalènes n'est pas sur le tronc de l'arbre, mais sur des branches horizontales élevées plus ou moins cachées. (On the selective forces acting in the industrial melanism of Biston and Oligia moths, K. Mikkola, Biological Journal of the Linnean Society, 1984, 21, 409; On Scientific Fakery and the Systems to Catch It, Kenneth Chang, The New York Times, October 15, 2002, D1; cités dans Exhuming the Peppered Mummy, J. Wells, 2007, August 30)
En conséquence, il n'y a aucune évidence qu'une sélection ait lieu de cette façon dans la nature.
Source: Phalène du bouleau et sélection naturelle, Gérard et Nadine Ciavatti
Dans le processus de sélection naturelle, les organismes les mieux adaptés à l'environnement survivent. Mais comment définit-on les mieux adaptés?
Des critiques ont montré que la réponse darwiniste est: ce sont les organismes qui ont la capacité de survivre (par exemple, Karl Popper, Science: problems, aims, responsibilities, 1963, Federation Proceedings 22, 961). Autrement dit, on tourne en rond, on ne fait que répéter la même chose. En philosophie c'est appelé une tautologie.
Étienne Rabaud (biologiste français, 1868 - 1956) écrit: L'hypothèse [darwiniste] ne résiste pas à la critique la plus élémentaire. Ne suffit-il pas de constater que l'appréciation d'un avantage tourne dans un cercle vicieux? Quand un organisme persiste, nous décidons qu'il possède une disposition avantageuse, et nous déclarons avantageuse une disposition quelconque, précisément parce que l'organisme persiste. (E. Rabaud, Introduction aux sciences biologiques, 1941)
Une critique identique plus récente a été développée par le biologiste et entomologiste français Rémy Chauvin (1913 - 2009). Pionnier par ses idées développées à partir d'expériences originales concernant la vie des abeilles, des fourmis et des oiseaux, il affirme que la notion d'adaptation est une tautologie (La biologie de l’Esprit, 1985; Dieu des fourmis, Dieu des étoiles, 1988; Le darwinisme ou la fin d’un mythe, 1997).
Sources: Étienne Rabaud dans wikipédia; Rémy Chauvin dans wikipedia; Un savant indocile, Rémy Chauvin, dans la revue 3e millénaire, juin 2010; Objections to evolution dans wikipedia; Does evolution qualify as a scientific principal?, Ariel Roth, Geoscience 1977 Origin 4, 1, 4
Pour justifier la théorie de la sélection naturelle et de l'ancêtre commun, les scientifiques s'appuient sur l'examen des organismes vivants et des fossiles.
Leur méthodologie consiste à trouver des similitudes dans les traits des organismes, nommées homologies, et à établir des correspondances entre elles. Puis ils repèrent des parentés probables et les représentent sous forme de diagrammes (arbres buissonnants).
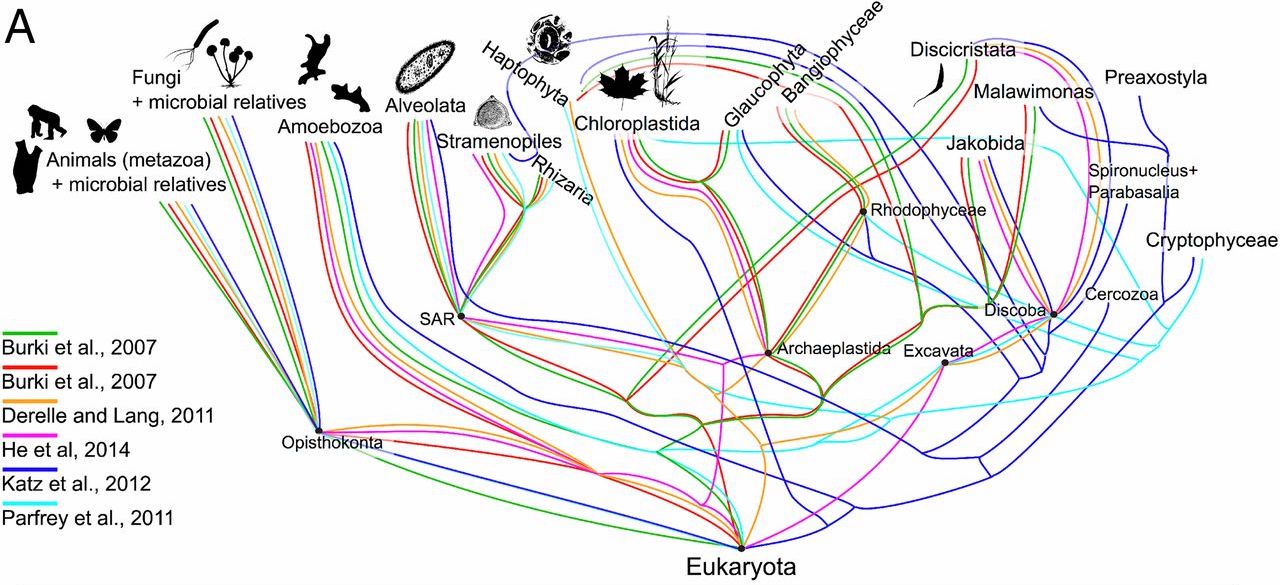
Proposition d'un "arbre de vie".
Extrait de Synthesis of phylogeny and taxonomy into a comprehensive tree of life, Cody E. Hinchliff et al., Proceedings of the National Academy of Sciences of the USA, 2015, 112, 41, 12764
On peut remarquer que certains animaux présentent des similitudes dans leur aspect morphologique, par exemple un trou dans la mâchoire.
On peut aussi remarquer des similitudes dans la disposition du squelette et des organes, leur organisation, et donc dans la structure globale de l'organisme. Par exemple, on observe le même plan d'organisation du squelette des membres chez le chat, la chauve-souris, le requin, l'oiseau et l'humain, qui sont tous des animaux vertébrés.
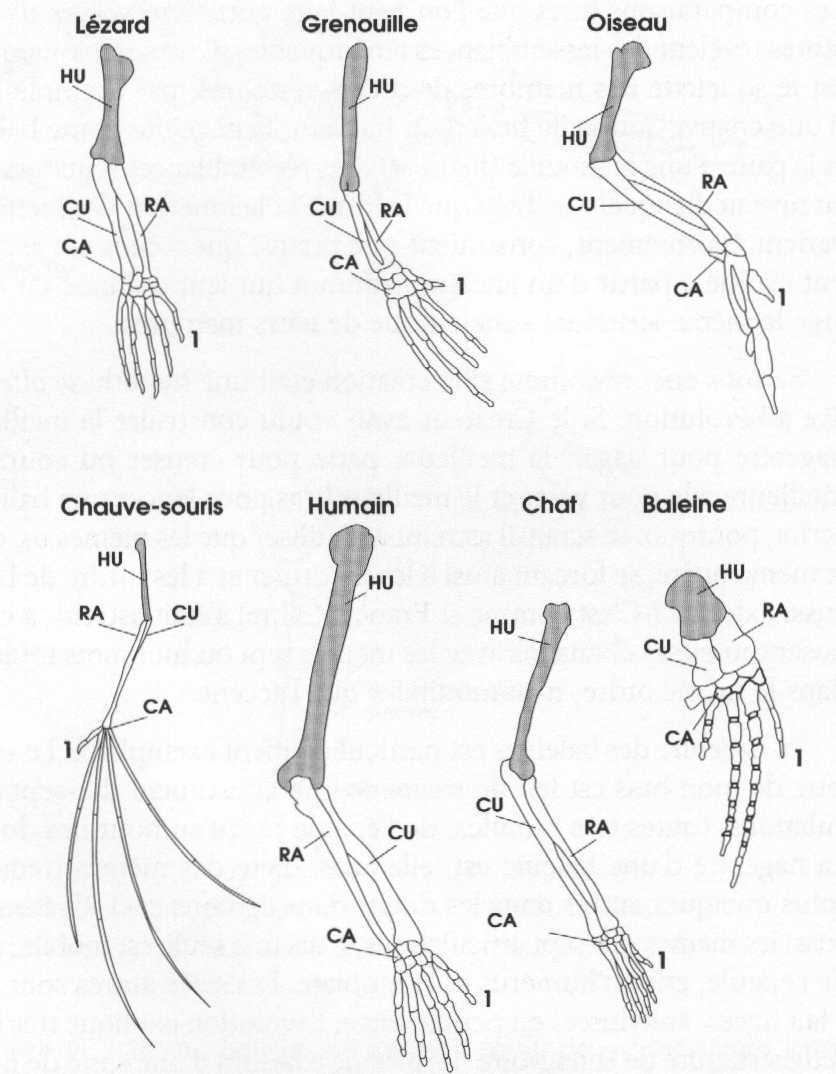
Squelettes comparés du membre antérieur de plusieurs vertébrés, sans tenir compte de leur taille. Les mêmes os sont ordonnés de la même façon. CA: carpe du poignet, CU: cubitus, HU: humérus, RA: radius, 1: pouce ou doigt numéro 1.
Merci à Les attributs des êtres vivants
Ces observations sont des faits concrets. Non seulement elles sont avérées, mais de plus ces homologies sont tout à fait remarquables.
On peut être émerveillé par l'organisation similaire de la nature pour les espèces considérées.
Les embryons constituent la forme à peine esquissée de l'individu adulte et pourtant, ils sont déjà bien structurés et leurs homologies sont frappantes. Par exemple les embryons des poissons, de la salamandre, de la tortue, du lapin, du poulet et de l'être humain sont très semblables.
Après la constatation des homologies, vient ensuite leur interprétation, forcément contrainte au cadre de la théorie néodarwiniste.
Un néodarwiniste considère que des caractères semblables dans deux espèces, qu'ils soient un détail morphologique ou un plan commun, sont le témoignage qu'elles dérivent d'un même ancêtre qui a évolué dans des lignées différentes.
Il en est de même pour les embryons. Leurs homologies seraient le témoignage que les animaux se sont développés à partir du même ancêtre. L'embryon montrerait le plan original de l'ancêtre.
Un archéologue qui découvre le squelette d'un animal inconnu va établir les homologies et différences avec d'autres squelettes connus, fossiles ou actuels.
L'exemple le plus fameux est celui de l'évolution du cheval depuis l'Eohippus (appelé anciennement hyracotherium), gros comme un chien, qui vivait au début de l'ère tertiaire il y a environ 50 millions d'années (voir chapitre précédent Histoire des animaux).
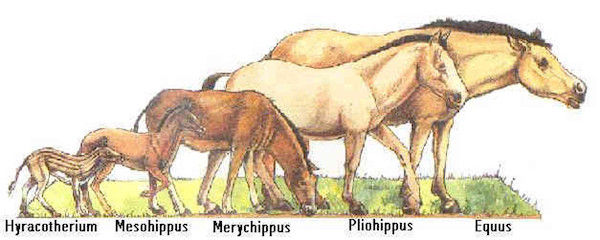
Animaux fossiles proches du cheval. Merci à Anfrix
Malheureusement, cet archéologue va sans délai interpréter les nouveaux caractères trouvés en terme d'avantage pour l'adaptation.
Selon le mathématicien français M-P Schützenberger (La Recherche 1996), les darwiniens disent que les chevaux ont augmenté leur taille pour s'enfuir plus vite et échapper aux prédateurs... On impose au lecteur l'idée que ce qui compte pour un ruminant, c'est la vitesse de fuite... Mais aucune raison biologique ne permet de déterminer si c'est le critère déterminant. Rien n'interdit de penser que l'augmentation de taille puisse avoir un effet négatif.
C'est un principe posé à priori qui n'est pas forcément justifié. Le principe d'un ancêtre commun est donc seulement une hypothèse de travail. Mais elle est devenue omniprésente et on a oublié son caractère d'hypothèse.
D'autres explications sont possibles, par exemple celle d'archétypes communs que j'exposerai dans la partie 4.
Si l'on maintient malgré tout l'interprétation unique d'un ancêtre commun, on en fait une croyance au lieu d'une hypothèse. Et lorsqu'on assimile cette croyance à un fait avéré - ce qui est fait couramment dans les textes scientifiques et les média (voir partie 3) - cela tourne à la tromperie.
Ainsi, nombreuses sont les similitudes constatées dans des organismes qui ne sont pas reliés de près. C'est le cas par exemple pour des parties spécifiques du corps. Elles ne peuvent pas être imputées à un ancêtre commun.
Ainsi la nageoire du dauphin (un mammifère) et celle du requin (un poisson) sont semblables bien qu'ils ne soient pas de proches parents dans l'arbre évolutif proposé. Des similitudes existent aussi entre les ailes de la mouche et celles des oiseaux.
Dans ce cas, la similitude n'est pas appelée homologie, mais analogie, ce qui est seulement un changement de terminologie de surface. La cause supposée de l'analogie est que les organismes ont adopté la même structure parce qu'ils vivent dans des environnements semblables, remplissent une fonction spécifique et ont la même façon de s'adapter en développant le même plan.
C'est ce que les néodarwinistes nomment la convergence. C'est un autre type de mécanisme évolutif dans le monde darwinien. Cela semble plausible.
Mais alors, comment prouver que les homologies constatées ne sont pas tout bonnement des analogies dues à des convergences? Il semble qu'on les définisse comme homologies simplement parce que cela conforte la théorie de l'ancêtre commun.
Ainsi A.J. Jones écrit: L'argument évolutionniste de l'homologie manque de contenu scientifique. Ce manque particulier a de très sérieuses implications; il attaque la racine de toutes les tentatives par les évolutionnistes de donner à l'homologie une base objective et de distinguer homologie (similitudes dues à la descendance) de analogie (similitudes non dues à la descendance)... On reconnait maintenant que le concept évolutionniste d'homologie est entièrement subjectif. (A.J. Jones, A creationist critique of homology, CRSQ, 1981, 19, 3, 156, cité dans Does homology provide evidence of evolutionary naturalism? J. Bergman, Journal of Creation, 2001, 15, 1, 26)
Les évolutionnistes pensent que les organes et parties anatomiques dont ils n'ont pas découvert de fonction sont inutiles. Ils les considèrent comme des vestiges de structures qui ont été utiles à des formes ancestrales, puis sont devenues inutiles dans les formes plus évoluées, mais sont demeurés dans le corps parce que la nature les a oubliés.
Les biologistes considèrent comme vestiges chez l'être humain: les poils, la dent de sagesse, l'appendice, le coccyx (une queue atrophiée), les mamelons des hommes. Pourtant, plus la connaissance avance et plus des organes classés comme vestiges se sont révélés avoir une fonction utile. Il est bien présomptueux de penser qu'un organe est inutile. Nous sommes loin d'avoir découvert toutes les fonctions et mécanismes biochimiques et électriques du corps.
Par exemple, on sait depuis 2007 seulement que l'appendice vermiculaire est un organe lymphoïde qui contient des cellules immunitaires productrice d'anticorps et qu'il contribue à l'équilibre de la flore intestinale. Les thérapeutes psychocorporels, les ostéopathes, les naturopathes et les énergéticiens savent que les poils ont une fonction réceptrice, à la manière d'une antenne, sans parler de leur rôle dans l'aération de la peau. Le coccyx, quant à lui, sert de point d'insertion de plusieurs muscles et ligaments. Il est associé au centre de notre énergie vitale qui manifeste les besoins de base (sentiment de sécurité, nourriture, abri sur la tête, protection, amour) (voir article Chakras et corps subtils).
Sources: La classification du vivant, mode d'emploi, Gilles Camus; Les preuves de l'évolution, MERveille-NATure, 2008
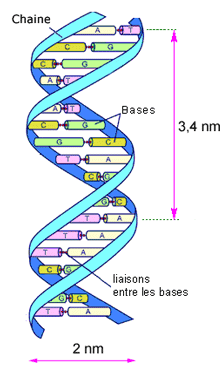
Structure de l'ADN: 2 chaines enroulées en hélice, reliées par leurs nucléotides opposés
L'approche moderne de l'évolution inclut le niveau moléculaire et l'ADN. Rappelons que l'ADN est constitué de 2 brins opposés, supports d'une suite de nucléotides (adénine, guanine, cytosine, thymine). Les deux brins sont reliés par leurs nucléotides opposés. Un gène est constitué d'un petit fragment de cette suite, donc d'une suite courte de nucléotides. (voir La molécule d'ADN et le code génétique)
D'un individu à l'autre, on peut repérer le même gène par son emplacement (locus) sur la molécule d'ADN. Mais la séquence des nucléotides de ce gène peut varier de quelques nucléotides d'un individu à l'autre, et d'une espèce à l'autre. Ces différentes versions d'un même gène sont nommées allèles (voir Wikipédia, allèle). Il peut donc exister dans les populations animales et humaines différentes allèles pour un même locus.
Il en est de même pour les protéines, qui sont constituées d'un enchainement d'acides aminés. Les séquences d'acides aminés d'une même protéine peuvent varier d'une espèce à une autre.
En comparant le génome de deux espèces, les généticiens repèrent les gènes communs (homologies) et les divergences. Ils partent du principe que les homologies sont le signe qu'il existe un ancêtre commun entre les espèces considérées. Les divergences (variétés d'allèles) auraient été provoquées par mutation, et auraient conduit à des espèces ayant évolué séparément.
Les biologistes relèvent aussi des homologies entre des gènes d'espèces différentes. Comment les relier aux homologies des traits anatomiques? On pourrait logiquement penser que des traits anatomiques homologues sont programmés par des gènes homologues.
Il n'en est rien. Il n'y a pas de correspondance entre les homologies des gènes et les homologies des structures qu'ils contrôlent. Des structures homologues de différentes espèces sont souvent produites par dès gènes complètement différents. Des organes qui apparaissent identiques chez différents animaux ne sont pas issus du même groupe de cellules de l'embryon.
Inversement, des gènes similaires peuvent contrôler des structures complètement différentes chez des espèces distinctes. Par exemple le même gène contrôle la taille chez la souris et la morphologie de l'organe sexuel chez la mouche drosophile.
Selon la théorie synthétique de l'évolution, les modifications génétiques (les allèles) se produisent de façon aléatoire et incontrôlée et sont transmises à la descendance. Il est important d'examiner quelle est la réalité de cette affirmation sur le plan biologique.
Au cours du temps, sous l'influence de l'environnement, il est attesté que des séquences de l'ADN ou de l'ARN peuvent être modifiées (voir La molécule d'ADN et le code génétique). Les facteurs de l'environnement qui provoquent ces modifications sont essentiellement des rayonnements électromagnétiques, des rayonnements de particules ionisées, ou des produits chimiques.
Sous l'influence de rayonnements ultraviolets par exemple, des liaisons entre atomes sur un brin de l'ADN peuvent être brisées et cela libère de petits groupements chimiques. Des liaisons entre nucléotides opposés peuvent aussi être brisées. En conséquence, des réactions chimiques peuvent se produire entre groupements voisins. Ainsi, la cytosine peut devenir de la thymine ou s'accoler à une autre thymine voisine. Cela crée donc des anomalies dans la structure de l'ADN, appelées mutations.
Dans le cas où ces anomalies ont lieu dans les gènes, cela modifie le code génétique et cela entraine des dysfonctionnements dans l'organisme. (Voir annexe Conséquences néfastes des mutations)
Lorsque la mutation affecte des cellules sexuelles (c'est-à-dire les cellules qui donnent naissance à des ovules ou des spermatozoïdes), les anomalies peuvent être transmises aux progénitures. La mutation est alors héréditaire.
Ces mutations sont très étudiées comme causes avérées de cancérisation.
On estime que l'ADN d'une cellule humaine moyenne subit plusieurs dizaines de milliers de lésions par cellule et par jour, dans des conditions habituelles d'activité métabolique et d'exposition aux facteurs environnementaux.
Toutefois, le corps est une merveilleuse machine extrêmement complexe, et on ne cesse d'en découvrir davantage les ressources astucieuses. Ces lésions restent normales et transitoires, car elles sont rapidement réparées.
Ainsi, lorsqu'une anomalie est détectée dans l'ADN (simple cassure, modification chimique, pontage entre deux parties de la molécule ou avec une autre molécule), une équipe de molécules - ouvriers spécialisés - intervient automatiquement afin que le corps conserve son intégrité. C'est un système très efficace.
La réparation consiste en une sorte de travail de couture pour enlever ce qui est en trop ou abimé, reconstruire ce qui manque et refaire les liens. Aussi, les mutations persistantes sont-elles extrêmement rares.
Le zèle employé par l'organisme pour conserver son intégrité n'est pas cohérent avec l'affirmation néodarwiniste que les forces naturelles tendent à produire des variations par mutation.
Des modifications dans les gènes peuvent parfois se produire au cours des processus de division des cellules.
Pendant cette période, l'ADN se dédouble et les gènes sont recopiés (duplication des gènes). Des erreurs de copies surviennent parfois lorsqu'un gène, ou une partie de gène, saute d'un endroit du génome à un autre, sur une partie codante ou non-codante, sur le même brin ou sur un autre. C'est le phénomène de transposition, bien documenté, qui provoque un brassage génétique.
Selon la théorie néodarwiniste, le gène copié continuerait à subir des mutations qui s'accumuleraient régulièrement, jusqu'à ce qu'une nouvelle fonction apparaisse.
La théorie néodarwiniste affirme que par ce mécanisme de recombinaison au cours de la duplication des gènes et de mutation que la vie aurait évolué depuis la bactérie, qui comporte quelques gènes primordiaux, jusqu'aux organismes supérieurs comportant des dizaines de milliers de gènes.
En conséquence, la théorie prédit une corrélation entre la complexité de l’organisme et la taille du génome. Mais cette corrélation ne se vérifie pas dans les faits. Ainsi les humains ont environ 30'000 gènes alors que le riz en a 50'000. L'épigénétique nous apportera un nouvel éclairage à ce sujet (voir plus loin).
La théorie néodarwiniste repose sur une chaine de mécanismes supposés, tels que les mutations aléatoires. Si un des chainons manque, cela met la théorie en défaut. L'un des chainons les plus contestés, et particulièrement étrange, est l'idée que des mutations aléatoires qui s'accumulent conduisent à l'apparition d'un nouveau gène qui procure des avantages à l'espèce.
Or des modifications de gènes sont extrêmement rares, et nous avons vu que lorsqu'elles se produisent, elles sont le plus souvent corrigées immédiatement. Si toutefois elles échappent à la correction, les mutations conduisent dans la presque totalité des cas connus à des dégradations plutôt qu'à des améliorations. (Voir annexe Conséquences néfastes des mutations)
Y aurait-il tout de même la possibilité que certaines mutations soient avantageuses pour l'espèce. Autrement dit certaines mutations aboutissent-elles à des gènes capables de produire une nouvelle protéine qui engendrerait un ou plusieurs caractères spécifiques supérieurs en terme d'adaptation chez l'individu?
Dans des scénarios que nous proposent certaines bandes dessinées et films de science-fiction, on voit agir des super-héros dotés de super-pouvoirs apparus grâce à une mutation génétique spéciale. Dans la BD X-Men, les héros sont capables de voler, de lire les pensées des autres ou encore de contrôler les champs magnétiques à la suite de mutations. Est-ce réaliste? Absolument pas.
Le chercheur français Denis Dupuy, chargé de recherche à l'INSERM, affirme: En général, une mutation génétique se fait rarement au bénéfice de l'individu. Concrètement, une mutation va plutôt engendrer une maladie que vous rendre capable de contrôler des tornades. (X-Men: La fiction pourrait-elle un jour devenir réalité? Le HuffPost, 21/05/2014)
Les biologistes pensent que des mutations peuvent arriver au hasard dans n'importe quel gène, n'importe quelle cellule, à n'importe quel moment. En conséquence, il est possible de leur appliquer les lois mathématiques de probabilité, et d'introduire des vitesses de mutation de fragments d'ADN.
On appelle fréquence de mutation le nombre de molécules biologiques (séquences de nucléotides de l'ADN ou séquences d'acides aminés des protéines) qui ont muté dans une population donnée (de la bactérie à l'homme).
Le taux de mutation est la fréquence de mutation survenue dans un laps de temps donné (un an, ou une génération, etc.). Il fournit la probabilité mathématique qu'un gène soit différent de celui de la cellule-mère. Pour le génome humain, les biologistes acceptent le nombre de 70 modifications par génome et par génération. Mais cela varie selon les organes ou tissus physiologiques envisagés.
Alors que ce n'est qu'une supposition, les scientifiques acceptent la régularité de mutation comme une base solide, au point qu'ils ont élaboré une technique de datation génétique nommée horloge moléculaire. En comptant les mutations détectées dans une espèce par rapport à une autre, ils en déduisent le temps passé depuis la séparation de ces deux espèces.
C'est de cette façon qu'ils ont énoncé que les Néandertaliens et les Humains se sont séparés il y a 600'000 à 750'000 ans.
En réalité, les taux d'évolution varient des procaryotes aux eucaryotes, d'une espèce à l'autre, d'une partie de l'ADN à une autre, et selon qu'on analyse les gènes ou les protéines. Ils varient aussi dans le temps, incluant des moments d'arrêt. Aussi, les estimations de cette horloge sont-elles souvent en désaccord avec la datation des terrains géologiques.
Des chercheurs ont tenté de calculer la probabilité pour qu'un nouveau caractère émerge par une accumulation de mutations conduisant à un nouveau gène.
Le Dr Douglas Axe a estimé la probabilité pour qu'une nouvelle séquence fonctionnelle de protéine se produise parmi un vaste ensemble de combinaisons possibles. Il a trouvé 1 chance sur 1077 (1 suivi de 77 zéros) - (Axe Douglas, Estimating the Prevalence of Protein Sequences Adopting Functional Enzyme Folds, Journal of Molecular Biology, 2004, 341, 1295). D'autres chercheurs sont arrivés à des conclusions semblables.
Le Dr Stephan Meyer (épistémologue états-unien, né en 1958, auteur de Darwin’s Doubt) en conclut que l'hypothèse qu'une recherche aléatoire ait réellement produit l'information génétique nécessaire à une innovation morphologique dans l'histoire de la vie est de façon écrasante bien plus susceptible d'être fausse que vraie. (Stephan C. Meyer, Dawkins’s Dilemma: Misrepresent the Mechanism…or Face the Math).
Le Dr John Sanford et son équipe ont développé un logiciel pour simuler l'évolution des populations par mutation génétique. Ils en ont conclu que la théorie génétique de l'évolution n'a aucun support théorique - c'est un modèle scientifique indéfendable. (Using Numerical Simulation to Test the Validity of Neo-Darwinian Theory, J. Sanford, J. Baumgardner, W. Brewer, W. ReMine, & P. Gibson, In Snelling Ed., 2008, Proceedings of the Sixth International Conference on Creationism, p.165)

Marcel-Paul Schützenberger
Merci au site M-P Schützenberger
Médecin et mathématicien français, élu membre de l’Académie des sciences en 1988, Marcel Paul Schützenberger (1920-1996) a joué un rôle déterminant dans la création de l'informatique théorique en France. Il est un pionnier de la théorie des langages, en collaboration avec le linguiste états-unien Noam Chomsky (né en 1928).
Sur la base de démonstrations mathématiques et de la théorie du langage, il a démontré que la fabrication d'objets biologiques merveilleusement compliqués et efficaces requiert une quantité considérable d'information qui est sans rapport avec le nombre d'instructions nouvelles qui peuvent être introduites par accumulation de mutations. Le hasard est donc incapable de produire quoi que ce soit de valeur.
Même si l'on peut comprendre que des espèces peuvent être défavorisées dans certaines circonstances environnementales et que certaines peuvent disparaitre, nos connaissances biologiques ne nous permettent pas de prédire qu'une espèce est favorisée par le milieu. La sélection naturelle n'explique pas grand-chose.
Sources: Principal
causes of hot spots for cytosine to thymine mutations at sites of cytosine methylation in growing cells, E. Lutsenko, A.S. Bhagwat, Mutation Research, 1999, 437, 1, 11; Mécanismes de réparation des dommages, Éric Quémeneur, CEA Visiatome, 2007; Réparation
de l'ADN, Cours de pharmacie, Matthieu Simon, 2008;
La duplication des gènes, moteur de l'évolution, Joseph Schacherer, La Recherche 01/04/2009, 429, 50; La
duplication de gènes fournit-elle le moteur de l’évolution? Jerry Bergman, Journal of Creation, 2006, 20, 1, 99
Évolution des taux de mutation dans Wikipédia; Rates
of Spontaneous Mutation, John W. Drake, Brian Charlesworth, Deborah Charlesworth and James F. Crow, Genetics, 1998, 148, 4, 1667; Genetic
drift, selection and the evolution of the mutation rate, Michael Lynch, Matthew S. Ackerman, Jean-Francois Gout, Hongan Long, Way Sung, W. Kelley Thomas & Patricia L. Foster, Nature Reviews Genetics, 2016, 17, 704; Horloge
moléculaire dans Futura-sciences; L'horloge moléculaire remise à l'heure? 2013, dans Hominidés; L’horloge moléculaire, qu’est-ce que c’est? Al Tanner, 2015, et Comment
les horloges moléculaires donnent l’heure de l’espèce humaine, B. Alex et P. Moorjani, 2018, dans The Conversation
Les failles du darwinisme, M-P Schützenberger, La Recherche, 283, janvier 1996 (english version The miracles of Darwinism);
Sité dédié à Marcel-Paul Schützenberger, avec en particulier toutes ses publications; Marcel-Paul
Schützenberger - French Darwin doubter, Jerry Bergman, Journal of Creation, 2014, 28, 2, 123
Dans le modèle darwiniste, le mécanisme de la mutation par les gènes est basé sur une certaine représentation de la structure et du fonctionnement de l'ADN centré exclusivement sur les gènes. Or notre connaissance du fonctionnement de l'ADN a beaucoup changé depuis quelques années, grâce aux apports des techniques fines de chimie moléculaire et d'analyse génétique.
L'un des apports récents majeurs de la génétique a été le séquençage de l'ADN. Le premier génome séquencé fut celui de l'homme en 2001. Puis suivirent ceux d'autres espèces animales (bactéries, drosophile, souris, lézard,... - voir en France le Génoscope du CEA)
Plus la recherche avance, plus le modèle de l'ADN et de son fonctionnement se précise et se complexifie, et plus les représentations antérieures se révèlent inadéquates.
Dans le modèle initial, l'ADN est fragmenté en gènes et en zones dites non-codantes. Le gène porte le code de fabrication d'une protéine spécifique identifiée, qui est transmis par l'intermédiaire d'un ARN (voir mon article L'ADN et le code génétique). Dans ce cadre de pensée rudimentaire très linéaire,
un gène = un ARN messager = une protéine = une fonction
Le modèle actuel nous montre un ADN dont les fragments ont des fonctions complexes et variables. Dans mon article l'ADN et ses modes d'expression, j'ai décrit le phénomène des gènes sauteurs ou transposons, ainsi que celui de l'épissage alternatif dans lequel un même gène peut coder pour des protéines différentes selon les parties qui sont rendues inactives (introns).
D'autres phénomènes rendent le système de lecture encore plus élaboré et plus complexe, donc plus difficile à appréhender. Il s'agit en particulier du changement de cadre de lecture, et de la découverte que les zones non-codantes sont partiellement codantes.
Même sans compter l'épissage alternatif, pourquoi un gène correspondrait-il à une seule protéine? De façon très logique, un même gène peut être lu de 6 façons différentes et a donc le potentiel pour coder 6 protéines différentes.
En effet, le code se présente comme la succession de groupes de 3 nucléotides, appelés codons. La lecture commence donc à un nucléotide et procède de 3 en 3. Or, si le début est décalé d'un ou de deux nucléotides, on obtient un autre cadre et une toute autre lecture, comme le montre le schéma suivant.
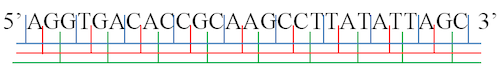
Exemple de lecture d'une chaine de nucléotides.
On commence à gauche, au chiffre 5. Puis on lit le premier codon de 3, AGG, et on continue de 3 en 3 (cadre bleu).
Mais il est aussi possible de commencer au premier G, et lire GGT (cadre rouge), ou de commencer au deuxième G et de lire GTA (cadre vert).
Merci à Wikimedia, travail personnel de Hornung Ákos
On a donc défini 3 cadres de lecture. Les 3 autres sont similaires, lorsque le code est lu sur le brin complémentaire dans l'autre sens (à partir de la droite - chiffre 3 - sur la figure).
Si plusieurs cadres de lecture sont opérationnels simultanément, alors la même portion d'ADN comprend non pas un seul gène, mais 2 ou 3, voire plus.
Il y a même la possibilité que le début de lecture d'un gène commence dans le corps d'un autre gène et se prolonge au-delà, soit des décalages supérieurs à 3. Ces gènes sont dits chevauchants.
Si une mutation survient, alors elle peut entrainer des conséquences sur plusieurs gènes chevauchants.
Les zones autres que les gènes ont été appelées non-codantes parce qu'elles ne portent apparemment pas de code de fabrication de protéines et qu'on ne voyait pas au début quel pouvait être leur rôle. Un moment, elles furent même appelées zones poubelles. (voir mon article Architecture et structure de l'ADN)
Or certaines parties de ces zones dites non-codantes ressemblent à des gènes, qui semblent défectueux ou inactifs. Aussi d'aucuns les considèrent comme des vestiges inutiles de l'évolution. Ils considèrent qu'ils ont été autrefois d'abord altérés par mutation, puis oubliés par la sélection naturelle, au même titre que les organes taxés de vestigiaux. Ils les nomment gènes fossiles ou pseudogènes.
Or on sait maintenant que les pseudogènes peuvent avoir une fonction.
Dans une expérimentation sur des souris, une équipe de scientifiques japonais a montré que la perte de l'un de ces pseudogènes entrainait la mort des souris étudiées. Le pseudogène avait bien un rôle, non reconnu avant. C'est celui de produire un microARN qui contrôle l'expression d'un autre gène vital (équipe de Shinji Hirotsune, Saitama Medical School, The Gems of "Junk" DNA, W.W. Gibbs, Scientific American, nov 2003).
D'autres recherches confirment et étendent ce résultat. En décryptant le génome complet d'une souris commune, une équipe américaine et britannique de chercheurs a montré en 2002 que les zones non-codantes sont riches en informations qui contrôlent l'activité des gènes. Les zones d'information seraient aussi étendues que les zones contenant les gènes.
Les pseudogènes fonctionnels sont fortement impliqués dans des taches variées - mise en silence de gènes, catalyse, régulation de croissance - et ils auraient donc une importance dans l'apparition de malformations et de maladies. En 2010, des équipes ont analysé le mécanisme de contrôle de ces pseudogènes en détail et mis en avant le rôle d'ARN spécifiques. (A coding-independent function of gene and pseudogene mRNAs regulates tumour biology, Laura Poliseno et al., Nature, 2010, 465, 1033)
Comme l'écrit Claire Peltier dans Futura Santé, Le dogme de la biologie vient d'être remis en question par un groupe de généticiens. Les pseudogènes, reliquats du passé, ne seraient pas aussi désuets que ce que l'on a cru jusqu'à présent.
La façon dont une zone est catégorisée comme codante ou non doit donc être révisée. Des pseudogènes actifs peuvent échapper à l'analyse. Les auteurs d'une étude de 2018 affirment qu'une multitude de gènes codants importants ont été manqués à cause de la terminologie de zone non-codante. Ils suggèrent que beaucoup d'autres gènes et de fonctions pourraient être découverts. (The translation of non-canonical open reading frames controls mucosal immunity, Ruaidhrí Jackson et al., Nature, 2018, 564, 434)
De mon point de vue, affirmer qu'une zone est inutile et n'est qu'un vestige de l'histoire alors qu'on sait peu de choses montre une certaine prétention. Il serait plus honnête et plus humble de dire qu'on ne sait pas. Avec ces nouvelles connaissances, le mécanisme de mutation génétique transmissible apparait simpliste.
Sources: Do any vestigial organs exist in humans? Jerry Bergman, 2000, Journal of Creation, 14, 2, 95; Cadre de lecture dans wikipedia; Décryptage du génome de la souris de laboratoire, Futura-Santé, ADIT, 19/12/2002; The Mouse Genome And The Measure of Man, NIH News, december 2002; A Genome-Wide Survey of Human Pseudogenes, David Torrents et al., 2003, Genome Res. 2003 Dec; 13(12): 2559–2567; Les pseudogènes vont-ils bouleverser la biologie ? Futura-Santé, Claire Peltier 25/06/2010; Scientists study genes misidentified as 'non-protein coding', Ziba Kashef, Yale University, Phys.org, 13/12/2018
Des études génétiques récentes ont démontré que le génome n'a pas la capacité par lui-même de déterminer les traits d'un individu. Il est seulement une mémoire, un stock de possibilités, qui peuvent être manifestées ou non. Il est semblable à un livre qui peut être lu ou non.
De même que les informations contenues dans un livre prennent vie lorsqu'on en prend connaissance, de même les gènes se manifestent seulement s'ils sont rendus actifs. Les zones non-codantes interviennent dans ce contrôle. La science qui étudie les mécanismes de cette activation est l'épigénétique (voir mon article L'ADN et ses modes d'expression)
On peut dresser une carte des gènes qui sont activés ou non. C'est la configuration épigénétique ou épigénome.
Le principal mécanisme biologique opérant est la méthylation de l'ADN. Un groupe méthyl (-CH3) est ajouté à des nucléotides. Il agit comme une coiffe qui vient mettre le gène en veilleuse. C'est surtout la cytosine qui subit cette méthylation. Dans un génome, il est possible de dresser la carte des endroits qui sont méthylés.
Une conséquence importante est que la configuration épigénétique est en réalité beaucoup plus importante que le génome lui-même. C'est grâce à elle par exemple que le même matériel génétique qui se trouve dans des œufs de papillon s'exprime à un moment sous la forme d'une chenille et à un autre sous celle d'un papillon.
Avec ces nouvelles données, le biologiste états-unien Michael Skinner (né en 1956) et son équipe ont réexaminé les pinsons des Galapagos et ils ont montré que les variations de caractéristiques étaient corrélées aux nombreuses variations épigénétiques. (Epigenetics and the Evolution of Darwin’s Finches, M.K. Skinner et al., Genome Biology and Evolution, 2014, 6, 8, 1972)
Or ce système de contrôle peut être modifié par la pression de facteurs environnementaux et par le vécu de l'organisme, y compris le vécu psychologique.
De plus, ces empreintes épigénétiques peuvent être transmises à la descendance sans qu'il y ait une modification du génome. Selon Skinner, une transmission épigénétique a été observée dans un grand nombre d'espèces, incluant des plantes, des insectes, des poissons, des oiseaux, des rongeurs, des porcs et des humains. (Unified theory of evolution, Michael Skinner, 09/11/2016)
C'est contraire aux représentations antérieures des généticiens dans lesquelles seules les mutations incluses dans les gènes peuvent transmettre des modifications de traits. L'accent mis sur les mutations génétiques, les erreurs de copie et la transmission génétique à la descendance perd de sa pertinence. En effet, les changements de caractéristiques d'un organisme apparaissent plus comme étant dus à des modifications épigénétiques.
Or une caractéristique essentielle d'une configuration épigénétique est qu'elle est réversible, aussi bien au cours de la vie d'un individu que dans sa descendance. Se pose alors la question de savoir si cette transmission est éphémère ou durable. Skinner répond que des variations de traits et de maladie ont été suivies pendant plus de 10 générations sur la plupart des organismes y compris des mammifères, plus de 50 pour des vers, et jusqu'à une centaine pour des plantes.
Aussi, les pressions environnementales effectives, telles que des variations de température, d'ensoleillement, de ressources nutritionnelles, de composition chimique de l'atmosphère, peuvent induire rapidement des changements de traits et de nouvelles adaptations animales, sans avoir à passer par la mutation génétique et son aspect aléatoire d'une durée excessive.
Le mécanisme de sélection naturelle perd de son sens puisque tous les organismes ont la capacité de répondre à la pression environnementale.
Skinner ajoute: Je suis convaincu que nous avons atteint un point où un changement de paradigme est nécessaire.
Sources: Why everything you've been told about evolution is wrong, Oliver Burkeman, The Gardian, 19 mars 2010; A Comeback for Lamarckian Evolution? Two new studies show that the effects of a mother’s early environment can be passed on to the next generation, Emily Singer, MIT Technical Review, February 4, 2009
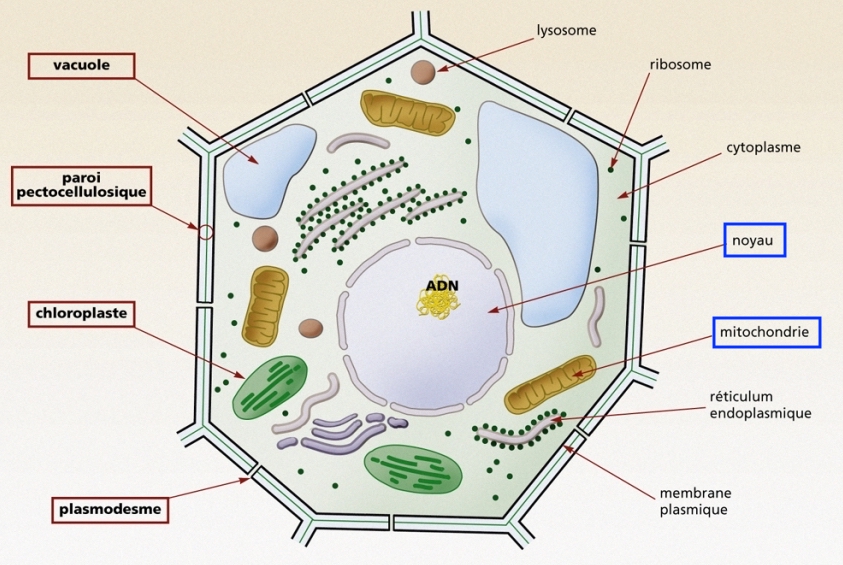
Schéma de la structure de la cellule végétale, avec le noyau et quelques mitochondries
Merci à GNIS Pédagogie
Depuis le début de cet article, lorsque nous évoquons les gènes et l'ADN d'un eucaryote, nous parlons de l'ADN qui se trouve dans le noyau. Le N de ADN signifie nucléaire parce que les molécules d'ADN ont été découvertes d'abord dans le noyau.
Par la suite on en a aussi trouvées dans les organites de la cellule appelées mitochondries, qui sont les centrales d'énergie de la cellule. Elles sont nombreuses, puisqu'on en dénombre environ 100 à 1000 par cellules. Chez l'humain, chaque mitochondrie contient une dizaine d'exemplaires d'ADN.
L'ADN des mitochondries est différent de celui du noyau. Sa taille est beaucoup plus petite (chez l'humain 16'569 paires de nucléotides, à comparer aux 3,3 milliards de paires de nucléotides de l'ADN nucléaire). Sa forme est généralement circulaire (en particulier chez l'humain), mais ce n'est pas toujours le cas.
Comme l'ADN nucléaire, l'ADN mitochondrial comporte des parties qui portent le code de fabrication de protéines, qui sont donc des gènes. Mais ils sont beaucoup moins nombreux, moins de 100. Chez l'humain, il y en a 37.
Enfin, l'ADN mitochondrial se transmet par la mère. Les mitochondries paternelles sont détruites au moment de la fécondation de l'ovule. Les enfants héritent donc du même ADN mitochondrial maternel (bien qu'un cas de transmission par le père ait été signalé en décembre 2018 dans un cas pathologique - Biparental Inheritance of Mitochondrial DNA in Humans, Shiyu Luo et al., PNAS 2018, 115, 51, 13039)
Toutefois, il existe des erreurs de transcription, considérées comme des mutations. Dans la recherche darwiniste de la séparation des lignées d'espèces, le moment de mutation de l'ADN mitochondrial est utilisé comme point de divergence.
Les recherches du biologiste canadien Paul D.N. Hebert (né en 1947) l'ont mené à la conclusion que chaque espèce pouvait être identifiée de façon nette par un gène spécifique de son ADN mitochondrial, le COI, qui occupe environ 5% du génome.
De cette façon, il a pu entre autres identifier les espèces d'oiseaux de façon concluante, en accord avec ce qui était déjà connu. C'est pourquoi Hebert a nommé ce gène le code-barre de l'ADN. Plus de 100'000 espèces ont été ainsi enregistrées à ce jour.
Sources: Identification of Birds through DNA Barcodes, Paul D.N. Hebert, Mark Y Stoeckle, Tyler S Zemlak, Charles M Francis, PLOS biology, 28/09/2004 - Biological identifications through DNA barcodes, Hebert PD, Cywinska A, Ball SL, deWaard JR, Proc Biol Sci. 2003, 270, 1512, 313
Si les espèces dérivent progressivement l'une de l'autre par une dérive génétique comme l'affirment les darwinistes, on devrait constater que plus les espèces se répandent et se multiplient, plus leur carte génétique se diversifie.
Or, en utilisant l'identification mitochondriale, deux généticiens, Mark Stoeckle et David Thaler, l'un états-unien, l'autre suisse, ont montré que ce n'est pas le cas. La diversité génétique n'est pas plus large chez les 7,6 milliards d'humains que chez les 500 millions de moineaux ou les 100'000 bécasseaux. (Why should mitochondria define species? Mark Y. Stoeckle and David S. Thaler, Human Evolution, 2018, 33, 1, 1)
De plus, on devrait trouver dans les différentes espèces des génomes qui sont proches les uns des autres et qui se distancient de façon continue. Les conclusions des auteurs sont contraires à cette attente. Chaque espèce est génétiquement différente de celle des autres, et les frontières sont très nettes. Chaque espèce a sa séquence spécifique étroite.
On ne trouve pas d'espèces avec des séquences génétiques intermédiaires. Ce sont, disent-ils, des ilots définis dans un océan de possibilités de séquences.
Les auteurs ont été surpris par leurs résultats et par ses répercussions possibles dans l'esprit de la communauté scientifique, au point qu'ils se sont crus obligés d'ajouter un avertissement en préambule: Cette étude repose et renforce fortement l'évolution darwiniste, incluant la compréhension que toute vie dérive d'une origine biologique commune depuis plusieurs milliards d'années.
Cette note est très révélatrice des tensions qui surgissent dès qu'un résultat semble en contradiction avec la théorie admise (voir partie 3). Les auteurs veulent rassurer leurs collègues en affirmant qu'ils restent bien dans le cadre imparti. Et pourtant, les résultats de l'étude montrent que ce n'est pas le cas, car ils posent une vraie énigme aux tenants de l'évolution darwiniste.
Sources: ADN mitochondrial, site de la police scientifique; The Father of DNA Barcoding, Katherine Tweed, ScienceLine, 01/02/2008; ADN mitochondrial: les pères le transmettent aussi, Futura santé, Marie-Céline Ray, 29/11/2018; Dossier l'ADN mitochondrial, PodcastScience, 20/01/2011; Évolution, vraiment? Une étude génétique d’envergure révèle que 90 % des espèces animales sont apparues en même temps, Fawkes News, 08/06/2018; Far from special: Humanity's tiny DNA differences are 'average' in animal kingdom, Rockefeller University, Phys.org news, 21/05/2018
Ce qui vient d'être exposé montre clairement que les biosciences ont beaucoup évolué depuis la formulation de la théorie néodarwiniste des années 1940, puis 1970. Elles ont fourni une abondance de connaissances sur les facteurs biologiques susceptibles d'induire des changements dans les caractéristiques des animaux (leur phénotype).
Aussi, de nouveaux courants scientifiques sont nés pour proposer des théories qui intègrent ces nouvelles connaissances biochimiques dans une théorie de l'évolution. C'est par exemple le cas de la biologie évolutive du développement dite evo-devo, née dans les années 1980, qui part du constat que la relation entre le phénotype (les caractères visibles) et le génotype (les gènes de l'ADN) n'est pas une simple question de lecture de gènes, mais que des facteurs extérieurs au génome jouent un rôle important dans le développement d'un organisme.
Un autre mouvement, la synthèse évolutive étendue (SEE), se distance nettement du néodarwinisme en soulignant les différences. Elle s'est développée sur la base de critiques de la théorie darwiniste moderne, comme la théorie des équilibres ponctués de Stephen Jay Gould dans les années 1980. Elle a été conceptualisée en 2007 par le biologiste et philosophe italien Massimo Pigliucci (né en 1964) et par le biologiste autrichien Gerd B. Müller (né en 1953), qui ensemble ont édité un livre sur ce sujet Evolution: The Extended Synthesis (MIT press, 2010).
La SEE prend en compte la complexité du fonctionnement des cellules sur le plan moléculaire. Se détachant de la conception du tout provient du gène, elle s'efforce d'élaborer une vision qui englobe non seulement l'organisme entier, mais aussi la population et l'environnement écologique.
Dans son bel article de présentation Why an extended evolutionary synthesis is necessary (Interface Focus, 2017, 7, 5, disponible ici), Gerd Müller écrit: Le cadre de travail insiste sur le rôle des processus constructifs, des interactions écologiques et de la dynamique des systèmes dans l'évolution de la complexité organique aussi bien que des conditions sociales et culturelles. La causalité linéaire à un seul niveau est remplacée par la causalité réciproque à niveaux multiples... Le cadre étendu... entraine une compréhension renouvelée du rôle de la sélection naturelle dans le processus évolutionnaire.
La SEE inclue en particulier:
Il est maintenant reconnu que des formes de transmission non-génétique sont opératives: épigénétique, mais aussi comportementales, écologiques, et culturelles.
Cette voie a été explorée en particulier par la généticienne israélite Eva Jablonka (née en 1952) dans son ouvrage Evolution in Four Dimensions: Genetic, Epigenetic, Behavioral, and Symbolic Variation in the History (avec Marion Lamb, 2005)
La comparaison des génomes des différentes espèces anciennes nous a appris qu'au cours de l'évolution des portions significatives du génome ont été recopiés, supprimés, ou réorientées sur une autre fonction (en codant pour une autre protéine); que des segments peuvent sauter et provenir d'autres cellules ou d'autres organismes (transfert horizontal de gènes), et pas seulement de leurs ascendants, aussi bien chez les eucaryotes que chez les procaryotes (animaux primitifs comme les bactéries).
En conséquence, l'évolution ne peut pas procéder par changement progressif et substitution d'allèles, mais répond par impulsion.
Tout en restant dans le cadre du principe de la sélection naturelle, des chercheurs ont proposé de corriger ses failles en étendant le principe de sélection au niveau du groupe. La compétition aurait lieu pas seulement entre individus dans un groupe, mais entre groupes pour le bénéfice du groupe.
À partir de 2001, des chercheurs généralisèrent ce principe de sélection de groupe à une sélection à multiples niveaux, en-dessous et au-dessus de l'individu, depuis la cellule jusqu'à l'espèce.
Alors que le darwinisme stipule que les conditions environnementales sont déterminantes dans le processus de sélection - sous forme de pression environnementale, la SEE met en évidence les influences qui s'exercent dans le sens inverse: de l'organisme vers son environnement.
En vivant, en se nourrissant, en déféquant, en construisant leur habitat, en se déplaçant, parfois sur de grandes distances, etc., les populations d'organismes modifient leur environnement. Elles construisent des "niches", autrement dit un environnement écologique local. Elles sont actives dans l'élaboration de conditions de vie favorables pour elles et pour d'autres populations.
Depuis les travaux de mathématiques et de physique de pionniers de la pré-informatique comme Ludwig von Bertalanffy ou Alan Turing, il est clair qu'un système composé de nombreux éléments en interactions multiples est plus que la somme de ses éléments. Il est le produit des interactions entre ses éléments. C'est le cas des organismes, qui peuvent être considérés comme des systèmes biologiques.

Pr Denis Noble. Merci à D. Noble et Twitter
Cela conduit à élaborer des modèles biologiques qui rassemblent des échelles multiples, de la molécule aux populations. Les gènes en sont seulement une petite partie. Des processus d'auto-organisation y sont à l'œuvre, tels qu'ils sont décrits par la branche mathématique nommée dynamique du chaos (ou physique des phénomènes irréversibles). Certains changements du phénotype sont expliqués par ces processus internes d'auto-organisation.
Le biologiste britannique Denis Noble (né en 1936), fervent partisan de la SEE, ajoute que les métaphores employées dans le langage biologique, bien implantées dans la culture scientifique, telles que programme génétique, livre de la vie, ou encore gène égoïste, faussent notre vision de la vie par leur caractère hautement réductionniste. À cela, il oppose la musique de la vie dans laquelle l'organisme biologique, loin d'être l'esclave de ses gènes, est une combinaison harmonieuse de causes multiples – un orchestre sans chef d'orchestre qui prend appui sur la biologie des systèmes. Il est l'auteur de La musique de la vie: La biologie au-delà du génome (Seuil, 2007) et de Dance to the Tune of Life (2016).
Sources: Entretien
avec le mathématicien Marcel-Paul Schützenberger: les failles du darwinisme. La Recherche 283, janvier 1996, disponible ICI;
Why everything you've been told about evolution is wrong, The Guardian, mars 2010; Does
evolutionary theory need a rethink? Kevin Laland et al., 2014, Nature, 514, 7521;
dans Wikipedia, Biologie évolutive du développement et Extended
evolutionary synthesis; Group selection; Niche
construction;
Evolution beyond neo-Darwinism: a new conceptual framework, Denis Noble, 2015, Journal of Experimental Biology, 218, 7, doi: 10.1242/jeb.106310, disponible ICI
Dans L'origine des espèces (p.200), Darwin a écrit: Si l’on arrivait à démontrer qu’il existe un organe complexe qui n’ait pas pu se former par une série de nombreuses modifications graduelles et légères, ma théorie ne pourrait certes plus se défendre.
Eh bien, c'est justement ce qui est arrivé.
Plus les biologistes explorent les mécanismes moléculaires de la vie et découvrent les détails microscopiques de leurs structures et de leurs rouages, plus ils mettent en évidence des systèmes complexes dont le fonctionnement implique la synergie de nombreux éléments interdépendants.
C'est par exemple le cas du flagelle des bactéries. Les bactéries sont munies d'une sorte de queue, le flagelle, qui leur permet de se propulser (voir Histoire des animaux). Il tourne comme l'hélice d'un bateau à la vitesse de 40 à 60 tours/s. Le mouvement de rotation se produit grâce à un mécanisme très astucieux qui comporte plusieurs pièces, dont un stator, un rotor et un embrayage, toutes réalisées avec des protéines.
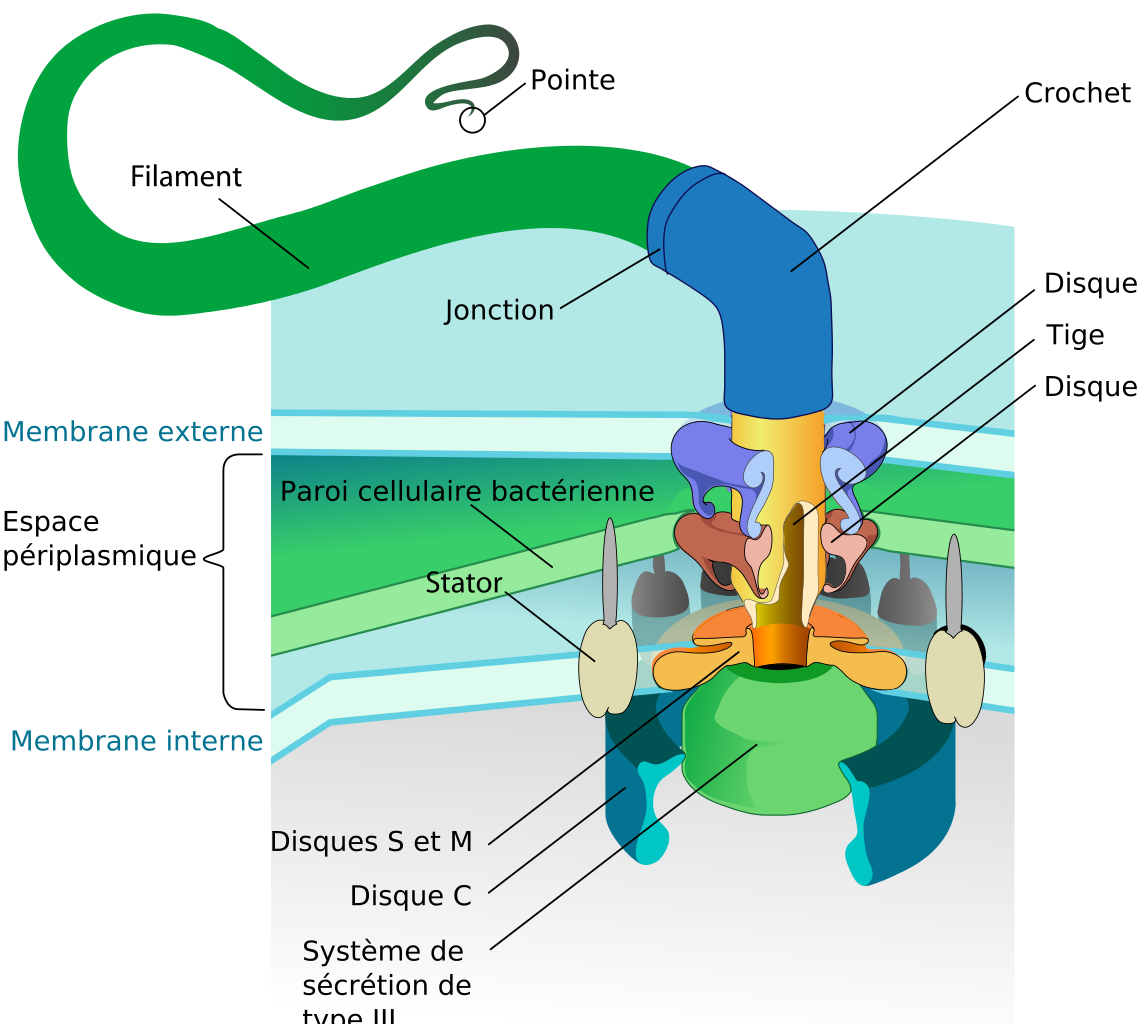 | 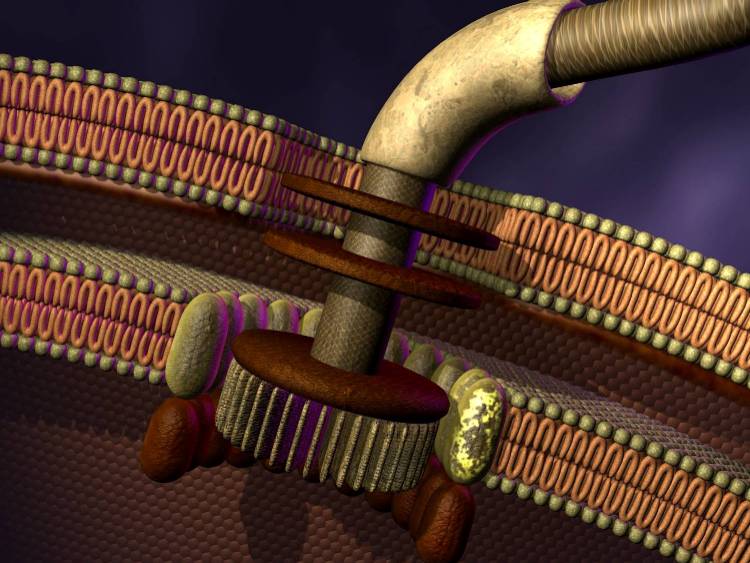 |
| Flagelle d'une bactérie Gram-négative | Représentation du mécanisme du flagelle. Merci à Evolution News |
Le plus étonnant est que la fabrication et le fonctionnement de ce moteur requièrent de nombreuses réactions biochimiques toutes synchronisées. Par exemple, la synthèse du flagelle met en œuvre l'expression orchestrée et synchronisée de plus de 60 produits géniques.
Or de tels systèmes apparaissent tout faits avec l'espèce bactérienne. Il ne peut pas y avoir eu de constructions rudimentaires préalables où les éléments auraient été ajoutés un à un. Si le système est incomplet, s'il manque une seule pièce, il ne fonctionne pas et n'apporte aucun avantage sélectif à l'espèce.

Pr. Michael Behe. Merci à Lehigh University
C'est ce que Michael Behe (né en 1952), professeur de biochimie états-unien à l'Université de Lehigh en Pennsylvanie, nomme la complexité irréductible. Elle qualifie un système composé de plusieurs parties qui interagissent, et qui ne peut plus fonctionner dès lors qu'on enlève n'importe laquelle de ces parties.
Michael Behe a publié ses recherches sur les failles de la théorie néodarwiniste dans un premier livre paru en 1996, Darwin's Black Box: The Biochemical Challenge to Evolution (traduction française La Boite Noire de Darwin: L'Intelligent Design, Presses de la Renaissance, 2009).
D'autres machines moléculaires naturelles ont été mises en évidence dans les organismes, servant au transport de protéines ou d'électrons, à la photosynthèse, et à beaucoup d'autres fonctions. Des chercheurs de l'université de Harvard (USA) ont découvert l'existence d'un moteur rotatif moléculaire responsable du glissement sur une surface de verre, d'une bactérie dépourvue de flagelle et de cils. Remarquons que toute mutation génétique briserait les gènes qui pilotent la fabrication de ce moteur.

Toutes ces machines ne peuvent pas avoir d'intermédiaires qui auraient progressivement évolué vers leur fonctionnement biochimique complexe actuel, comme le voudrait la théorie darwinienne. Behe donne l'analogie d'un piège à souris. La fonction de ce système ne se manifeste que lorsque le système est complet. Il ne peut pas y avoir une progression d'une espèce à l'autre dans l'assemblage de ces éléments par une progression aveugle.
On peut aussi comparer l'apparition d'un système complexe biologique tout constitué à l'invention de la roue. La roue ne peut fonctionner que si elle est parfaite. Il n'y a pas eu d'évolution étape par étape. Si sélection naturelle il y a eu éventuellement sur ce genre de systèmes moléculaires, elle n'a pu agir que sur des formations complètes.
Sources: Evidence for Intelligent Design from Biochemistry, Michael Behe, Discovery Institute, 1996; Michael Behe Hasn’t Been Refuted on the Flagellum, Evolution News, 2011; A Third Rotary Motor Has Now Been Found in Bacteria, Evolution news, 2015 et Current biology, 2015
Avant Behe, plusieurs auteurs ont exposé leurs doutes ou leur désaccord avec la théorie darwiniste. Parce qu'ils sont souvent passés sous silence, je tiens à en faire connaitre quelques-uns.
J'ai déjà présenté Fred Hoyle dans mon article Apparition de la Vie, astronome britannique (1915 - 2001), auteur de l'expression big bang, inventeur de la théorie de la panspermie ou vie terrestre provenant de microorganismes de l'espace.
En 1983, il publia un livre The Intelligent Universe, a new view of creation and evolution. Il y fustige l'idée du hasard présidant à l'apparition de la vie dans la soupe primitive. C'est dans ce livre que sont présentés des arguments devenus populaires: improbabilité qu'une construction moléculaire complexe se produise par hasard, improbabilité que des singes tapant au hasard sur une machine à écrire puissent composer un texte de Shakespeare. Il avance l'idée que les mutations sont incapables de conduire à une évolution, car elles ne produisent pas d'avantages. Il préfère l'hypothèse de la panspermie.
Il pense que l'apparition et l'évolution de la vie sont contrôlées par "une intelligence cosmique".
Michael Denton (né en 1943) est un biochimiste et auteur australien. Il est l'auteur du livre Évolution: Une théorie en crise (Flammarion, 1993), qui est une critique de la théorie synthétique de l'évolution. Il est paru en anglais en 1985, soit 2 ans après le livre de Fred Hoyle. Il y développe des arguments différents.
Ce livre a suscité des débats passionnés dans de nombreux pays. C'est lui qui a mis Michael Behe et d'autres sur la piste de l'examen des failles de la théorie darwinienne, et qui a motivé la naissance du mouvement de la Conception intelligente dont je parlerai dans la partie 4.
Denton admet la réalité de la sélection naturelle dans le cas des microévolutions (diversification au niveau de l'individu) comme l'allongement des becs de pinson (théorie mise à mal récemment comme je l'ai rapporté). Mais il n'y a aucune preuve tangible de macroévolutions (c'est-à-dire la diversification des grandes lignées) à partir d'un ancêtre commun. La complexité du monde biologique ne peut pas être mise sur le compte d'une extrapolation des microévolutions.
Les structures complexes ne peuvent pas être le résultat graduel de la sélection darwinienne. La sélection peut opérer pour conserver une structure globale mais pas pour la faire apparaitre. Denton renchérit dans son deuxième ouvrage, L'évolution a-t-elle un sens? (1997) en introduisant les processus moléculaires génétiques.
Milton rapporte que de nombreuses observations de faits scientifiques dans diverses disciplines ne sont pas en accord avec la théorie. Il dénonce le manque de preuves. Des similitudes entre espèces d'époques différentes n'indiquent pas forcément qu'elles dérivent l'une de l'autre. La variabilité ne fait pas apparaitre de nouvelles structures, mais seulement des variations légères. Des mutations favorables sont improbables.
Il montre aussi comment cette théorie est devenue un dogme.

Pr Didier Raoult
Merci à PACA-INSERM
Le biologiste Didier Raoult (né en 1952), est médecin de formation, spécialiste des maladies infectieuses. Dans son livre, Dépasser Darwin (2010, Plon), Didier Raoult explique pourquoi le darwinisme, érigé en dogme, est en train de voler en éclats, par suite des nouvelles compréhensions que nous avons sur la vie des microbes. Ils sont partout, y compris dans nos gènes, et nous cohabitons. (voir son interview dans Le Point, Et si Darwin s'était trompé, 12/12/2011)
D'autres auteurs mériteraient d'être mentionnés. Par exemple l'embryologiste danois Soren Lovtrup (1922–2002), auteur en 1987 de Darwinism: The Refutation of a Myth. Certains, comme Rémy Chauvin, sont présentés dans le fil de ces articles.
Sources: Fred Hoyle's The Intelligent Universe; A summary and review, Gert Korthof; Michael Denton dans wikipedia; Søren
Løvtrup dans wikipedia; Darwinism: The Refutation of a Myth, de Soren Lovtrup, analysé par Gert Korthof; Richard Milton dans wikipédia (anglais);
Shattering the Myths of Darwinism, disponible ICI
Un principe organisateur
Pratiques inquiétantes dans la science
3. Certains scientifiques et philosophes osent exprimer leur conviction de l'existence d'un principe organisateur supérieur qui guiderait les créations des organismes vivants. Le prix à payer est de faire la séparation dans leur esprit entre un domaine subjectif qui relève de leur foi et un domaine scientifique occupé par la théorie de l'évolution. Les chercheurs qui avancent des modèles scientifiques différents sont censurés, ridiculisés, ou évincés des institutions, bien loin d'un dialogue honnête qu'on pourrait attendre de la part de scientifiques éduqués. Alors qu'elle affirme abusivement être un fait objectif, la théorie synthétique de l'évolution est en réalité une hypothèse fondée sur des présupposés métaphysiques.
***
Lorsque la réparation ne parvient pas à restaurer la structure normale de l'ADN, quelles sont les conséquences de l'anomalie?
Le code génétique est une instruction pour la fabrication des protéines. Elle est analogue à un texte et une mutation est analogue à un changement dans les lettres de ce texte: suppression, addition ou déplacement. Sa conséquence dépendra de la fonction de la lettre modifiée et de son emplacement.
Voici une excellente illustration des types de mutation et de leurs conséquences par analogie avec des erreurs dans la copie d'un texte (extrait d'un cours de génétique de l'Université en ligne):
| Séquence de référence | On ne fait pas d'omelette sans casser des œufs | On comprend le sens de la phrase (le produit du gène est fonctionnel) |
|---|---|---|
| Mutation | Séquence modifiée | Conséquences de la mutation |
| Substitution | On ne sait pas d'omelette sans casser des œufs | Changement de séquence sans gravité |
| Substitution | On ne fait pas d'omelette sans masser des œufs. On ne fait pas d'omelette sans easser des œufs. | Faux sens Non sens |
| Délétion (suppression) | On ne fas pas d'omelette sans casser des œufs. On ne fait pas d'omelete sans casser des œufs. | Non sens Sans conséquence sur le sens |
| Addition | On ne fait pas d'omelette sans casser des bœufs. On ne fait pas d'ommelette sans casser des œufs. | Faux sens Sans conséquence sur le sens |
| Addition d'un STOP | On ne fait pas. // d'omelette sans casser des œufs | Non sens et fin de lecture anticipée |
| Duplication | On ne fait pait pas d'omelette sans casser des œufs. | Non sens |
| Déplacement | On ne fait pas d'œufs. | Non sens et fin de lecture anticipée |
| Échange | On ne fait pas d'œufs sans casser des omelette. | Non sens |
| Inversion | On ne fait pas d'ressac snas ettelemo des œufs. | Non sens |
Texte conforme à la nouvelle orthographe française (1990)
12 juin 2019




 Revenir à la première partie
Revenir à la première partie
{kind=link}
{kind=link}