
Une autre histoire de la Terre
2. L'histoire des animaux
racontée par les fossiles
Des bactéries à l'Être humain
Alain Boudet
Dr en Sciences Physiques
Résumé: Les fossiles exhumés des couches géologiques nous racontent comment les espèces animales ont successivement peuplé la Terre depuis les temps les plus anciens. On peut fournir un ordre chronologique des époques de leur vie, mais les datations sont des estimations incertaines. Sont d'abord apparus des organismes simples comme la bactérie. Puis se sont ajoutées des formes animales de plus en plus complexes, qui ont cohabité avec les formes simples, en équilibre écologique. Des analyses montrent que le peuplement et les extinctions se sont produits selon des rythmes cycliques. La cause la plus probable des extinctions serait une super-onde provoquée par l'explosion récurrente du noyau de la galaxie.
En examinant les fossiles, nous pouvons reconstituer une histoire approximative de la vie sur la Terre. Elle nous apparait comme la succession de nombreuses populations animales et végétales.
Dans le premier chapitre de cette série, j'ai décrit cette histoire depuis la formation de la Terre jusqu'à l'apparition des cyanobactéries (bactéries bleues, encore abondantes actuellement sur la planète). Vivant dans l'eau, elles ont produit de l'oxygène qui a été libéré dans l'atmosphère. Cela a créé un environnement favorable au développement de plantes et d'animaux terrestres, donnant une nouvelle orientation à l’évolution de notre planète il y a 2,2 milliards d'années (ou giga-années - Ga).
Dans ce deuxième chapitre, nous poursuivons la description de cette histoire jusqu'à l'apparition des êtres humains. Nous découvrons que des populations animales extraordinairement variées se sont succédées, non pas de façon continue et linéaire, mais par à-coups. Des populations ont disparu brusquement et d'autres sont apparues. Cela s'est produit selon un rythme cyclique.
Dans le chapitre suivant, nous examinerons les processus biologiques et informationnels susceptibles d'expliquer l'apparition de ces populations (théories de l'évolution).
Comment l'histoire des animaux peut-elle être reconstituée depuis l'origine de la vie sur Terre, alors qu'aucun témoin ne peut nous rapporter les scènes de vie de ces époques reculées?
Cela est possible uniquement à partir des traces que ces animaux ont laissées dans les roches sous forme de fossiles. C'est la découverte de ces fossiles qui a attiré l'attention de certaines personnes sur l'existence d'animaux ayant vécu dans des époques antérieures reculées. Certains ressemblent à des animaux vivant de nos jours, et d'autres non. Beaucoup appartiennent à des espèces disparues.
Les fossiles sont les restes minéraux d'un organisme, enrobés dans la roche. On en distingue deux sortes.
L'une contient des restes de l'animal: des parties minérales dures tels que les os, les dents, les carapaces ou les coquilles. Dans le cas des végétaux, ce sont les graines, les spores, le pollen.
Pour l'autre sorte, il ne reste rien de l'organisme qui a été complètement décomposé, surtout s'il s'agit de tissus mous. La trace qui subsiste est un moulage des tissus, leur empreinte.
Il existe un troisième type de traces, qui ne sont pas réellement un fossile, mais des traces chimiques. On les détecte en analysant la composition chimique de la roche. La difficulté est alors de savoir si elles proviennent vraiment d'un organisme ou si elles résultent de la transformation de la roche sous l'effet de changements climatiques et géologiques.
Le plus souvent, les organismes se décomposent sans laisser de traces. Il faut des circonstances favorables pour en laisser. Les animaux qui ont le plus de chance d'en laisser sont ceux qui sont composés de parties dures et ceux qui sont les plus abondants à une époque donnée. On estime que les espèces fossiles observées représentent moins de 6% du nombre d'espèces probables.
Les fossiles étaient déjà observés dans l'antiquité grecque, mais on ne savait pas ce qu'ils représentaient. C'est seulement à partir du 17e siècle et surtout au 18e que l'on a envisagé que des animaux et des végétaux marins avaient été enfouis dans des sédiments et ainsi préservés. Au 19e siècle, Charles Lyell (géologue britannique, 1797 - 1875), un ami de Darwin que nous retrouverons dans le chapitre suivant, émit l'idée que les fossiles sont des restes d'organismes ayant vécu à des époques antérieures.
De nos jours, environ 300'000 espèces de fossiles ont déjà été identifiées, des plus microscopiques aux plus volumineux.
La paléontologie est la discipline scientifique qui étudie les organismes fossilisés pour reconstituer la vie des temps géologiques.
En présence d'un fossile nouvellement découvert, le paléontologue se charge d'identifier l'animal ou la plante, ce qui n'a rien d'évident.
Ensuite, il tente de dater l'époque à laquelle la fossilisation a eu lieu en fonction de la couche (ou strate) de terrain dans laquelle le fossile est conservé. Enfin il consulte les données de datation qui ont été attribuées à cette strate, selon la méthode de stratigraphie du terrain.
La stratigraphie est née de la constatation que le terrain est fait de couches empilées plus ou moins bien marquées.
 |
| Couches géologiques bien marquées dans un terrain en Argentine, dans la région de Salta. Photo travelwayoflife. Merci à Wikimedia |
Les couches ont été formées lentement au cours du temps par des alluvions qui se sont déposées au fond de la mer ou d'un lac, ou encore qui ont été emportées puis déposées par une rivière ou par les vents. Parce que ce processus se répète à différentes périodes, plusieurs strates de composition différente se superposent. Les strates les plus profondes sont donc les plus anciennes.
Si on réalise une coupe dans ces sols, par exemple dans une carrière ou au moment de la construction d'une route, l'empilement des strates devient visible. La nature a elle-même réalisé des coupes dans les falaises, ou sur le flanc de montagnes, ou de terrains inclinés nommées affleurements. Ils sont le résultat de bouleversements géologiques qui ont incliné, plissé, fracturé ou soulevé les sols, de sorte que des sédiments marins peuvent se retrouver en altitude.
Dans chacune des strates sont inclus les fossiles de l'époque à laquelle elles se sont formées. Lorsque les géologues cherchent à les dater, ils appliquent le principe, pas toujours réalisé, que les couches les plus profondes sont les plus anciennes, et les plus élevées sont les plus récentes.
De plus, ils font appel à un principe issu de la paléontologie: deux couches ayant les mêmes fossiles caractéristiques ont le même âge. Il semble que certains types de fossiles répandus largement sur la Terre soient caractéristiques d'une époque géologique dans un temps limité. On les considère comme des fossiles indicateurs. Par exemple, les trilobites sont caractéristiques de l'ère paléozoïque et les ammonites le sont de l'ère mésozoïque (voir plus loin la définition de ces ères).
Le rapprochement des caractéristiques d'affleurements situés dans divers endroits de la planète ont conduit les paléontologues à jalonner la datation par des affleurements types.
Comme le principe d'âge égal des couches a une fiabilité limitée, les paléontologues utilisent de plus en plus des microfossiles comme indicateurs. On les trouve en abondance même dans des petits échantillons de sol. Par contre, on ne les voit qu'au microscope.
On trouve des fossiles dans les roches de tous les âges. Ce chapitre en présente un bref panorama pour chacune des différentes périodes géologiques.
Mais beaucoup ont été détruits parce que la roche a été transformée. On est donc très loin d'avoir un panorama complet de la succession des espèces animales au cours des millions d'années passées.
Nous constaterons que la succession des espèces n'est pas continue et tranquille. Elle a rencontré plusieurs bouleversements intenses. Dans la dernière partie de l'article, je présenterai des études qui montrent que ces bouleversements se sont produit selon des cycles.
L'examen des couches et leur mise en correspondance avec les fossiles ou microfossiles indicateurs conduit à leur attribuer un ordre de succession, c'est-à-dire à les classer chronologiquement l'une par rapport à l'autre (datation relative). Est-il possible de préciser l'époque à laquelle ces fossiles ont été enfouis et de leur attribuer une datation en années?
Il n'existe par de méthode directe pour cela. Aussi faut-il passer par des estimations indirectes. Ce sont des méthodes techniquement élaborées qui emploient la mesure de la quantité de produits radioactifs inclus dans les terrains étudiés, suivie d'un calcul théorique.
Or ce calcul repose sur des hypothèses et des principes qui ne sont pas systématiquement vérifiés, de sorte que les dates fournies sont des estimations incertaines.
Dans les couches géologiques, on détecte des éléments radioactifs qui sont inclus soit dans les minéraux eux-mêmes, soit dans les tissus organiques fossilisés.
Des atomes sont appelés radioactifs parce qu'ils sont instables et se décomposent en émettant des rayonnements.
Les rayonnements sont parfois des particules alpha, constituées de 2 protons et 2 neutrons (noyau d'hélium). Elles sont issues de l'élément radioactif, qui se transforme en un autre élément plus léger. Par exemple, l'uranium se transforme en plomb en émettant une particule alpha.
Si le rayonnement consiste en particules béta (électrons), l'atome radioactif se transforme en un autre atome de même masse atomique. Ainsi le carbone 14 se transforme en azote 14.
Toutes les méthodes de datation par éléments radioactifs fonctionnent sur le même principe. Le matériau géologique contient une certaine quantité d'atomes radioactifs au moment de sa formation, qui décroit au fur et à mesure que les atomes se désintègrent. Si on connait la quantité d'origine et la vitesse à laquelle elle décroit, on peut en déduire par le calcul le temps passé. Mais c'est justement ces éléments du calcul qui restent hypothétiques.
Les éléments radioactifs utilisés pour les datations sont principalement le carbone, l'uranium et le potassium. Le carbone est employé pour la datation d'objets organiques jusqu'à 50'000 ans au grand maximum. L'uranium est employé pour des minéraux ayant été formés il y a entre 10'000 ans et 1 Ga. Enfin le potassium fournit des dates pour des terrains volcaniques de 100'000 ans à 3 Ga.
La méthode de datation la plus connue du public est celle du carbone 14. Elle s’effectue uniquement sur du matériel qui provient d’organismes qui ont été vivants (bois, graines, pollen, ossements, cheveux, coquillages, insectes, etc.). Elle est basée sur les 2 principes suivants:
1. L'atmosphère contient du carbone ordinaire (carbone 12) sous forme de gaz carbonique et de méthane, ainsi qu'une petite proportion de carbone 14 radioactif. Ce dernier est produit en permanence par les rayons cosmiques à partir de l'azote. Un organisme vivant aérien s’imprègne de l’air qu’il respire et absorbe le carbone 14. On suppose que le rapport de la quantité de carbone 14 et de la quantité de carbone 12 (C14/C12) dans ses cellules est le même que celui de l’atmosphère qu’il respire.
2. Quand l’organisme meurt, il n’absorbe plus de carbone 14. Au cours du temps, le C14 de ses cellules se transforme en azote 14 par désintégration et le taux C14/C12 décroit lentement. On suppose que la décroissance se fait selon une loi mathématique basée sur une constante de temps caractéristique: la période de demi-vie. Cela signifie qu'au bout de cette période, la moitié des atomes se sont désintégrés. Pour le carbone 14, elle de 5730 ans.
Enfin, on mesure le taux actuel dans l'échantillon, et grâce à la loi mathématique, on en déduit le temps passé depuis le début de la décroissance, ce qui fournit l'âge de l'échantillon analysé.
Revenons sur le premier principe. On suppose que le rapport C14/C12 au moment de la mort de l’échantillon est celui de l'atmosphère de l'époque. Mais comment peut-on le connaitre, puisque personne n'était là pour le mesurer? On doit ajouter un troisième principe:
Les scientifiques considèrent que ce rapport a été constant au cours des temps dans l’atmosphère, et donc dans l’organisme, au moins depuis 50'000 ans, car les rayons cosmiques le régénèrent en permanence. On prend comme base les mesures effectuées en 1950.
Moyennant ces trois hypothèses, on peut donner une évaluation de l'âge d'un échantillon organique, tant que la quantité de C14 reste mesurable, soit 30'000 à 50'000 ans.
Revenons sur le deuxième principe: pourquoi la décroissance radioactive obéit-elle à une loi mathématique? Cette loi est basée elle-même sur deux suppositions.
1. On ne sait pas à quel moment un atome de C14 choisit de se transformer en azote 14. Les scientifiques sont muets sur ce sujet et ne cherchent pas les causes possibles. Aussi, dans une population d'atomes radioactifs, on suppose qu'un atome se désintègre au hasard. Cela peut lui arriver à tout moment. On traduit donc ce phénomène en termes de probabilités.
2. On suppose que la probabilité qu'un atome se désintègre est la même quel que soit le moment. C'est le taux de désintégration, et il est constant. Plus il est grand, plus la décroissance de la population est rapide, et de ce taux découle donc le temps de demi-vie. Mathématiquement, cela implique que la décroissance est exponentielle.
Connaissant le rapport actuel de C14/C12 fourni par la mesure de l'échantillon, ayant estimé la valeur de ce rapport au moment de la mort de l’organisme, l'application de la formule mathématique fournit le temps écoulé depuis la mort de l'organisme.
Deux atomes sont dits isotopes d'un même élément chimique s'ils ont le même nombre de protons, caractéristique cet élément, mais un nombre différent de neutrons. Dans la croute terrestre, l'uranium est relativement abondant sous la forme de deux isotopes, tous deux radioactifs.
L'un, l'uranium 238 se désintègre lentement en plusieurs étapes, se transformant en thorium, puis en plomb 206, en émettant une particule alpha. Sa demi-vie est de 4,5 Ga.
L'autre, l'uranium 235 se transforme en plomb 207, avec une demi-vie de 700 Ma environ.
Avec l'uranium, on n'a pas affaire à des organismes qui meurent, mais à des roches qui se sont formées après avoir été triturées, chauffées, pressées, etc.
Ici le principe est de supposer que la décroissance radioactive commence au moment où les atomes ont été emprisonnés par la cristallisation du minéral au moment de la formation de la roche (voir mon article Cristaux et géométrie cristalline).
Mais comment évaluer la quantité d'uranium présente au moment de la formation du minéral? On retrouve une difficulté semblable à celle de la datation au carbone 14.
À ce moment-là, on suppose que le plomb n'est pas encore présent et qu'il se forme au fur et à mesure de la désintégration de l'uranium. Cette supposition est discutable. La quantité initiale de plomb n'est probablement pas nulle à l'origine.
Enfin, pendant la décroissance, y a-t-il eu seulement la cause interne de décomposition selon la loi mathématique? Le système minéral n'a-t-il pas été influencé par des causes extérieures? S'il subit une hausse de température, il se décristallise et des éléments peuvent diffuser. Même chose s'il subit des pressions et des déformations. Enfin, l'uranium est soluble dans l'eau et peut avoir été partiellement lessivé.
À cause de ces incertitudes, seuls sont analysés des minéraux qui semblent se prêter plus facilement à des estimations, comme le zircon.
Avec ces estimations, en mesurant le rapport actuel U238/Pb206 dans l'échantillon, on peut calculer le temps écoulé grâce à la loi de décroissance. Mais on peut aussi mesurer le rapport actuel U235/Pb207 et calculer également un temps écoulé. Or les âges trouvés ne coïncident pas. Il faut donc comparer des terrains de même origine et appliquer des règles empiriques d'ajustement.
Une autre méthode bien employée en paléontologie est la mesure du rapport entre potassium 40, et son produit de désintégration, l'argon 40. Elle est employée pour des minéraux formés au cours d'une éruption volcanique.
La mesure de la quantité d'argon présente de nouvelles difficultés, car l'argon est un gaz qui peut s'évaporer, surtout si la roche a été soumise à des températures supérieures à 125°C. Dans ce cas, le temps calculé sera celui du dernier épisode d'exposition à la chaleur, non celui de la formation de la roche. De plus la roche peut avoir absorbé de l'argon présent dans l'atmosphère, ce qui rend la mesure imprécise.
Revenons à l'hypothèse du taux de désintégration constant et de la décroissance exponentielle supposée de tout produit radioactif. Certaines observations expérimentales la mettent en défaut.
Cette loi résulte d'une observation empirique limitée, traduite sous forme statistique. Elle permet de calculer des quantités moyennes, mais elle ne donne pas d'explications sur la cause de la désintégration d'un atome à un moment donné. Quels mécanismes sous-atomiques physiques sont à l'œuvre? Pourquoi ce moment et pas un autre? Les scientifiques ne proposent pas de solution.
Pourtant, un scientifique russe, le professeur Simon Shnoll (né en 1930) et son équipe de l’Institut de biophysique théorique et expérimentale de Pushchino (université d’État de Moscou) ont étudié expérimentalement cette question et ont obtenu des éléments de réponse. J'ai relaté les détails de ces expériences dans mon article Physique quantique, les concepts fondamentaux.
Pour Shnoll, une loi statistique est seulement une façon de masquer notre ignorance sur ces mécanismes. À partir de 1954, lui et son équipe ont inlassablement et minutieusement recueilli des données sur des processus réputés aléatoires en radioactivité, en biologie et en chimie. Ils ont constaté que tous ces processus obéissaient à des cycles de temps de 24 h, de 27,28 jours et de 365 jours. Or 27,28 jours est la durée de la révolution synodique du soleil et les autres cycles sont bien connus. Ils en ont conclu que la désintégration des atomes est influencée par le mouvement de la Terre et des planètes.
Nous pouvons en déduire que la décroissance n'est pas aléatoire, mais influencée par des événements cosmiques et qu'elle a pu dévier de la loi mathématique.
Malgré les incertitudes sur les datations normalisées, nous allons les considérer comme des points de repère commodes et intéressants, mais toujours sujets à une révision possible.
Sources: Principe de la méthode de datation par chaines radioactives, Institut Français de l'Éducation; Géochronologie, cours de l'université de Lille; Géochronologie U-Pb et microtectonique, Christian Pin, thèse 1979; Physics methods in Art and Archaeology, Institute for Structure and Nuclear Astrophysics, USA; Realization of discrete states during fluctuations in macroscopic processes, S.E. Shnoll et al, Uspekhi Fisicheskikh Nauk, 1998, 41, 10, 1025-1035)
De la découverte de milliers de fossiles sur tous les continents dans des strates de tous âges, de leur observation, leur étude et leur classification, se dégage un film: celui de l'évolution du peuplement animal. La Terre a été le théâtre de vie de nombreuses espèces animales.
Ceci est un fait incontestable. Il est tout à fait possible d'établir un ordre chronologique approximatif de ces entrées en scène et de reconstituer dans ses grandes lignes le film de la succession des familles animales.
Cela admis, la question qui vient à l'esprit est celle du mécanisme qui préside à cette évolution. De quelle façon, par l'action de quelles forces, les espèces apparaissent-elles, puis disparaissent-elles?
Imaginer un mécanisme qui fait apparaitre une espèce relève de la théorie. C'est un modèle explicatif possible, et cette démarche est normale en science. Penser par exemple que parce que deux espèces ont des structures semblables, l'une est dérivée de l'autre ou qu'elles dérivent toutes deux d'une troisième, n'est pas un fait, mais une hypothèse.
Le chapitre suivant décrira quelques causes plausibles de l'évolution de la vie sur la Terre.
En premier lieu, j'y décrirai la théorie de l'évolution de Darwin et de ses successeurs. Elle stipule que les espèces dérivent d'un ancêtre par mutations génétiques et sélection naturelle. Nous constaterons que c'est une spéculation qui comporte de nombreuses lacunes et qu'elle est vivement controversée. Il existe d'autres façons plus satisfaisantes d'envisager l'apparition des différentes espèces.
Dans ce site web qui, à la réalité matérielle et ondulatoire étudiée scientifiquement, associe d'autres réalités bien attestées des mondes supérieurs, il est évident qu'une théorie de l'évolution doit intégrer l'influence de champs d'information et de programmes provenant de plans d'intelligence supérieure. (voir mon article Au-delà de la matière: Les plans subtils d'intelligence supérieure. Une réalité tangible et incontournable)
Dans le présent chapitre, nous assisterons à l'entrée en scène chronologique des principales familles animales sur le théâtre de la Terre.
Je vous invite simplement à admirer la variété et la beauté de la Vie. Bon voyage.
Dans le chapitre précédent, nous avons assisté à l'apparition de la vie il y a 3,8 milliards d'années (ou 3800 millions d'années ou Ma) sous forme de bactéries primitives.
Les bactéries sont les organismes les plus simples que l'on connaisse. Elles sont constituées d'une seule cellule (organisme unicellulaire ou monocellulaire).
Des organismes plus complexes que les bactéries sont apparus, peut-être plus tard qu'elles, ou peut-être en même temps. Ils ont coexisté, comme je le préciserai en fin d'article. Leur complexité s'affiche à deux niveaux.
Ou bien, la cellule unique se structure intérieurement de façon plus élaborée.
Ou bien plusieurs cellules s'associent pour former un organisme (organisme multicellulaire ou pluricellulaire). Généralement, ces cellules se différencient et se spécialisent en organes.
Pour comprendre cela, examinons la diversité cellulaire des organismes actuels qui sont proches de ces organismes anciens.
Les bactéries choisissent parfois de se regrouper en amas sous forme de boules, de rubans ou de filaments. Ce sont des colonies de bactéries, comme c'est le cas des cyanobactéries observées dans le chapitre précédent. Il peut exister une forme d'organisation, un peu à la manière des populations de fourmis par exemple.
À propos de cette organisation, il est indiqué de mentionner un phénomène que les paléontologues ne prennent pas en compte. Depuis longtemps on sait que les organismes vivants, y compris les cellules bactériennes, peuvent communiquer entre elles et envoyer des informations par rayonnements électromagnétiques. Cette connaissance est le résultat de milliers d'expériences effectuées depuis les années 1930 (voir article L'ADN électromagnétique). Les informations transmises peuvent inclure des indications pour que la masse des cellules se comporte de façon coordonnée.
La plupart des plantes et les animaux qui nous sont familiers, ainsi que les êtres humains, sont des organismes composés d'innombrables cellules. C'est un assemblage de cellules spécialisées qui assument des fonctions différentes. Dans ce cas, il y a une véritable organisation interne de l'organisme.
L'être humain en est un exemple typique, avec des organes comme les dents, les yeux, l'estomac, la peau, etc., qui sont solidaires et tous indispensables.
Penchons-nous maintenant sur la structure interne d'une cellule de bactérie. On y trouve de nombreuses petites machines moléculaires élaborées.
La cellule ressemble à un sac qui enferme une substance gélatineuse (le cytoplasme). Dans cette substance, flottent l'ADN en forme de boucle, des enzymes (molécules de protéines) nécessaires à sa nutrition et à sa reproduction et d'autres assemblages moléculaires (voir figure dans le tableau ci-dessous). De la paroi, sort un cil (le flagelle) qui sert au déplacement de la bactérie de la même façon qu'une hélice sur le moteur d'un bateau.
Nous avons donc affaire à un organisme de structure simple. Mais attention aux mots. Cela ne signifie pas qu'il s'agit d'un organisme rudimentaire. La bactérie est un organisme complet et organisé qui est doté de toutes les fonctions de la vie: se nourrir, excréter, se mouvoir et se reproduire.
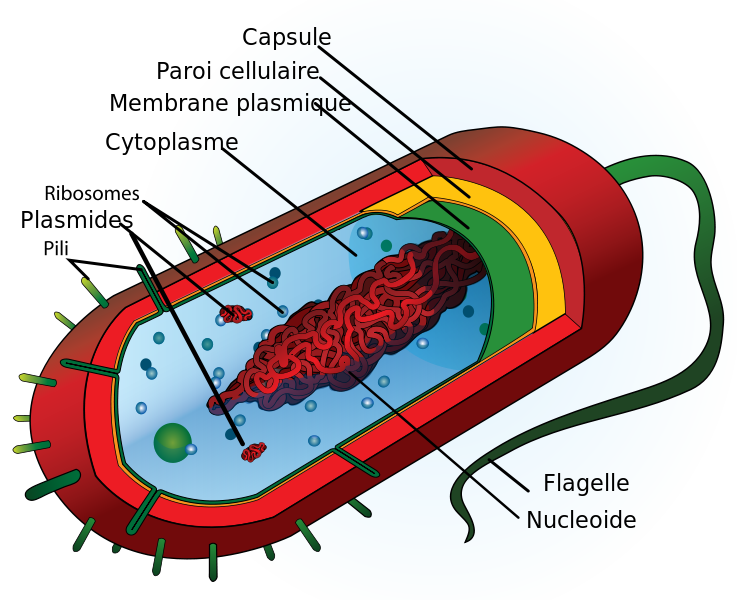 | 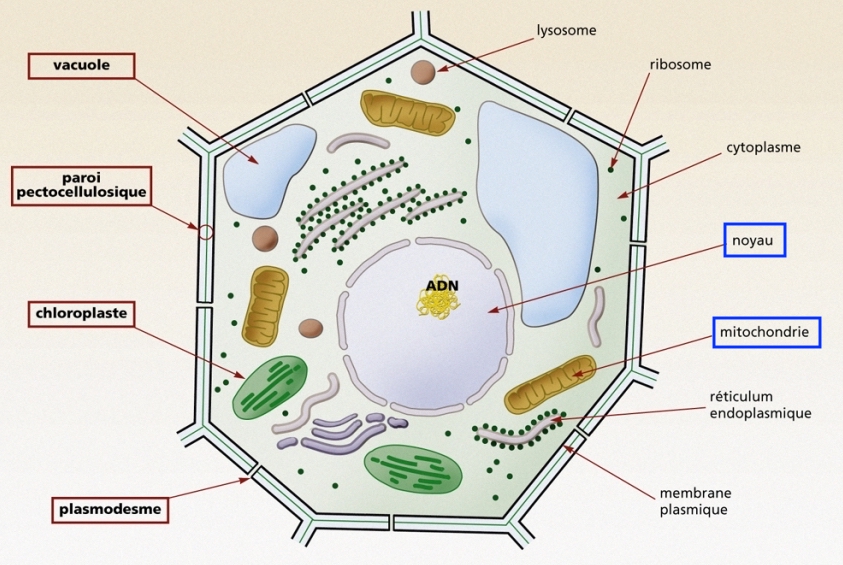 |
| Schéma de la structure cellulaire d’une bactérie (procaryote). Dessin de Mariana Ruiz Villarreal. Merci à Wikimedia | Schéma de la structure d'une cellule végétale (eucaryote). Merci à GNIS
Pédagogie |
Dans d'autres organismes, unicellulaires ou pluricellulaires, les cellules sont structurées de façon plus complexe. Le matériel génétique (l'ADN et les protéines qui forment les chromosomes) est séparé du reste du cytoplasme dans une vésicule appelée le noyau (voir article L'ADN moléculaire).
Les organismes composés de cellules avec noyau sont nommés eucaryotes (=avec un vrai noyau).
Par contraste, les organismes composés de cellules sans noyau comme les bactéries sont nommés procaryotes (=avant le noyau).
Les procaryotes ont tendance à être unicellulaires, mais nous avons vu qu'il existe aussi des colonies.
La très grande majorité des plantes et des animaux sont des eucaryotes multicellulaires.
Il existe toutefois des eucaryotes unicellulaires (nommés aussi protistes) tels que l'amibe et la paramécie.
Les champignons sont également des eucaryotes. Ni animaux ni vraiment plantes, ils sont reconnus depuis 1969 comme étant un règne par eux-mêmes, les mycètes ou fungi. C'est un groupe très large comprenant aussi bien des organismes unicellulaires microscopiques comme les levures, que des organismes pluricellulaires allant d'une taille microscopique (les moisissures) jusqu'aux champignons communs de nos forêts.
À l'intérieur des cellules à noyau (eucaryotes), des petites structures nommées organites ont des fonctions vitales. Ainsi, la mitochondrie, présente dans la majorité des espèces, joue un rôle important dans le production d'énergie. Un autre, le chloroplaste, est essentiel dans les végétaux qui utilisent la lumière pour synthétiser leurs molécules (photosynthèse). La cellule inclut également un système de filaments rigides formant une sorte de charpente, le cytosquelette. La cellule végétale se distingue de la cellule animale par la paroi cellulosique qui l'entoure (voir figure dans le tableau ci-dessus).
Les paléontologues ont découvert de nombreux fossiles d'organismes unicellulaires dès les années 1950. Les distinguer dans la roche n'est pas aisé, à la fois parce qu'ils sont très petits et parce que le processus aboutissant à la fossilisation de ces organismes mous est incertain.
En 1953, après les trouvailles de quelques pionniers, Stanley A. Tyler (géologue états-unien, 1906 - 1963) découvrit des micro-fossiles d'organismes unicellulaires datés d'environ 2000 Ma dans des stromatolithes dans le gisement de Gunflint dans l'Ontario au Canada. En forme de petites sphères, de bâtonnets et de filaments de moins de 10 µm, ils ressemblent à des moisissures ou à des algues.
Quelques années plus tard, à partir de 1965, J. William Schopf (paléontologue états-unien, né en 1941) étudia des fossiles très bien conservés remontant à 850 Ma environ dans le gisement de Bitter Spring dans le bassin du lac Amédée en Australie. De nombreuses espèces furent identifiées: cyanobactéries (procaryotes) sous forme de sphères et de filaments, acritarches (voir ci-dessous), mycètes et algues.
Bitter Springs a aussi livré la preuve convaincante de l'existence dès cette époque (850 Ma) de cellules eucaryotes d'algues vertes. Cette collection de micro-fossiles très anciens démontra qu'ils présentaient une grande variété.
Certains organismes unicellulaires à coquille trouvés à Bitter Spring peuvent difficilement être classés dans une catégorie connue. Cela leur a valu leur nom: acritarche provient de deux mots grecs signifiant origine incertaine, inclassable. On trouve des acritarches en abondance dans des gisements de moins de 1000 Ma. Mais ils étaient présents bien avant. On en trouve aussi dans des gisements de 1300 Ma dans le sud de la Chine (Province de Shanxi, Ruyang Group, gisement de Beidajian) et dans des strates de 1800 Ma dans le territoire russe.
Leurs formes sont très variées: sphère, étoile, fuseau, prisme. Ils sont formés d'une coque lisse ou dotée d'épines ou de cornes. Ils sont vraisemblablement eucaryotes, car leur unique cellule peut atteindre une taille importante de 10 à 80 micromètres (1 µm = 1/1000e de millimètre).
Sources: Occurrence of Structurally Preserved Plants in Pre-Cambrian Rocks of the Canadian Shield, Tyler Stanley A., Barghoorn Elso S., 1954, Science 119, 606; Microorganisms
from the Gunflint Chert, Barghoorn Elso S. and Tyler Stanley A., 1965, Science, 147, 3658, 563; Significance
of the Gunflint (Precambrian) Microflora, Cloud Preston E., 1965, Science, 148, 3666, 27
Microflora of the Bitter Springs Formation, Late Precambrian, Central Australia, J. William Schopf, 1968, Journal of Paleontology 42, 651
Solution to Darwin's dilemma: Discovery of the missing Precambrian record of life, J. William Schopf, 2000, PNAS, 97, 13, 6947; Cradle
of Life: The Discovery of Earth's Earliest Fossils, 1999, Princeton Univ. Press.
Protists of the Upper Mesoproterozoic Ruyang Group in Shanxi Province, China, Yin Leiming, Yuan Xunlai, Meng Fanwei, Hu Jie, 2005, Precambrian Research, 141, 1–2, 49
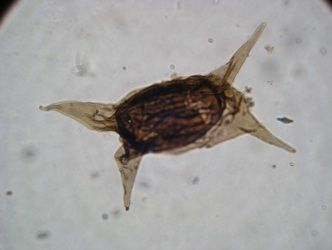 | 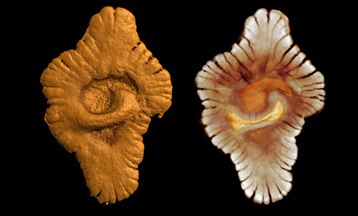 |
| Un acritarche | Un des fossiles, probablement multicellulaires, âgés de 2100 Ma, découverts en 2008 à Franceville au Gabon. À gauche, aspect externe, à droite aspect interne. Image reconstruite par microtomographie X. Photo © CNRS Photothèque / A. El Albani & A. Mazurier |
Des trouvailles successives plus récentes ont permit de reculer la date d'apparition des micro-organismes.
Dans le chapitre précédent, nous avons vu que les fossiles de micro-organismes vivants les plus anciens (des cyanobactéries procaryotes) ont été récoltés en mai 2017 dans un terrain australien, l'Apex Chert près de Pilbara, daté de 3500 Ma. La fossilisation a pu avoir lieu parce que la bactérie produit une coque autour d'elle.
Vers 1990, des centaines de fossiles de tubes et de rubans spiralés de 2 mm de diamètre et 1 à 50 cm de long, ont été découverts dans la mine de fer Empire, près de Negaunee dans le Michigan (46°27'N, 87°38"O USA), datée de 2100 Ma. Les auteurs les ont identifiés à Grypania spiralis, une algue rouge fossile qui a d'abord été découverte dans des sédiments plus jeunes (1100 Ma) en Chine et en Inde. Elle est considérée comme un eucaryote pluricellulaire.
En 2008, plus de 400 fossiles de formes régulières et de dimensions diverses ont été découverts au Gabon près de Franceville et étudiés par une équipe internationale de chercheurs coordonnée par A. El Albani du CNRS de Poitiers. Leur taille atteignant 10 à 12 centimètres montre qu'il s'agit de véritables organismes pluricellulaires marins inclus dans des sédiments datés de 2100 Ma.
En 2017, une équipe de paléontologues ont publié leur découverte de centaines de filaments fossiles de champignons (donc eucaryotes) dans le gisement Ongeluk en Afrique du Sud, province de Northern Cape, datés de 2400 Ma. Ce sont donc provisoirement les plus anciens eucaryotes.
Les populations de ces micro-organismes (bactéries, algues rouges ou vertes, champignons...) ont longtemps été les seules à peupler la Terre.
Et puis, vers -600 Ma, de nombreuses espèces d'animaux multicellulaires apparaissent et se diversifient, profitant d'une hausse subite de la concentration en oxygène dans l'atmosphère, d'abord avec la faune d'Ediacara, puis vers -540 Ma avec la faune de Burgess. Cela marque l'arrivée d'une nouvelle ère, l'ère paléozoïque, qui commence par la période du Cambrien.
Sources: Megascopic eukaryotic algae from the 2.1-billion-year-old negaunee iron-formation, Michigan TM Han et B Runnegar, 1992, Science, 257, 5067, 232;
A Morphological and Geochemical Investigation of Grypania spiralis: Implications for Early Earth Evolution, 2010, thèse de Miles Anthony Henderson, University of Tennessee, Knoxville
Paleobiology of the late Mesoproterozoic (ca. 1200 Ma) Hunting Formation, Somerset Island, arctic Canada, Nicholas J. Butterfield, 2001, Precambrian Research, 111, 1-4, 235
The controversial “Cambrian” fossils of the Vindhyan are real but more than a billion years older, Stefan Bengtson, Veneta Belivanova, Birger Rasmussen and Martin Whitehouse, Proc. Nat. Acad. Sci. USA, 2009, 106, 19, 7729
Découverte de l'existence d'une vie complexe et pluricellulaire datant de plus de deux milliards d'années, Communiqué du CNRS, 30 juin 2010; Ancient macrofossils unearthed in West Africa, Nature News, 30 juin 2010; Large colonial organisms with coordinated growth in oxygenated environments 2.1 Gyr ago, El Albani et coll., Nature, 2010, 466, 100
Anatomy and plant affinity of Chuaria, Wang Xin, Yuan XunLai, Zhou ChuanMing, Du KaiHe & Gong Miao, Chinese Science Bulletin, 2011, 56, 12, 1256
Paleobiological Perspectives on Early Eukaryotic Evolution, Andrew H. Knoll, 2014 Cold Spring Harbor Laboratory Press
Fungus-like mycelial fossils in 2.4-billion-year-old vesicular basalt, Stefan Bengtson et al., 2017, Nature Ecology & Evolution 1, 0141; Fossils
may be earliest known multicellular life: study, Marlowe Hood, site de nouvelles scientifiques, phys.org, 24 avril 2017
Au début de cet article, j'ai écrit que les affleurements et gisements archéologiques avaient été datés par comparaison avec des gisements types.
Puisqu'un tel classement inclut toujours une part de choix arbitraire, il a fait l'objet d'une normalisation par la Commission internationale de stratigraphie (ICS) et l’Union internationale des sciences géologiques (UISG), sous la forme d'un diagramme international de chronostatigraphie. Selon cette convention, les ères géologiques ont été définies par des dates de début et de fin, et ont reçu un nom officiel. Les grandes ères sont elles-mêmes divisées en périodes et époques.
Depuis l'apparition de la vie vers -4000 Ma jusqu'à -540 Ma, c'est l'ère du Précambrien.
Lui succède la période du Cambrien (de -540 à -485 Ma) qui marque le début de l'ère paléozoïque (ou ère primaire selon une terminologie plus ancienne). La racine "zo" dans protérozoïque et paléozoïque et dans les noms des ères suivantes fait référence aux animaux. La définition des différentes périodes est rapportée dans le tableau ci-dessous.
Sources: International Commission on Stratigraphy; Charte stratigraphique internationale
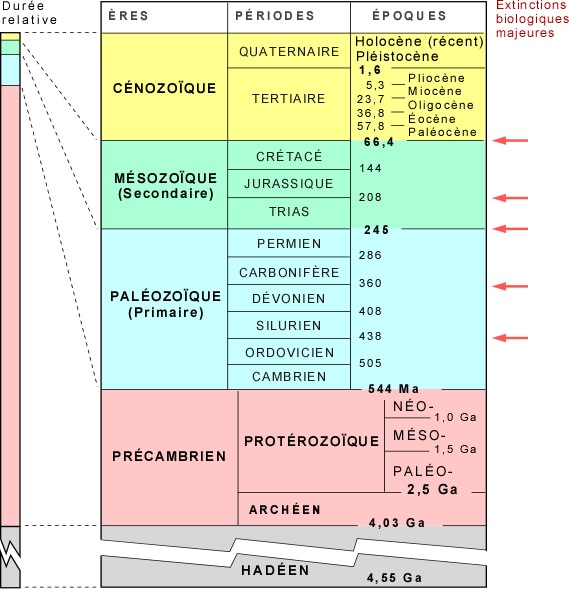 |
| Calendrier des ères et périodes géologiques (en bas les périodes les plus anciennes). Y sont indiquées les grandes extinctions d'espèces. Merci à Planète Terre (Pierre-André Bourque) |
Arrêtons-nous un instant dans notre récit de la succession des espèces animales pour récapituler leurs apparitions, en les situant comparativement dans le temps. Pour cela, nous reprenons et complétons l'image de l'horloge de 12h, proposée dans le chapitre précédent. 0 marque la formation de la planète et 12h représente notre présent. La durée de 12h représente donc 4,56 Ga.

Avec cette échelle, nous trouvons (en gras, ce qui est reporté sur le cadran)
Avant d'entrer dans le Cambrien, nous mettons doucement fin au Précambrien avec la faune d'Ediacara qui représente une pulsation soudaine de changement survenue il y a environ 600 Ma.
Ce sont essentiellement des organismes à corps mous, donc sans squelette, dont on ne trouve que les empreintes. Ils ont des formes de disque, de coupe, de tube, de feuille, etc. Ils ressemblent à des méduses, des vers, des coraux, ou à rien de connu. Leur taille varie de quelques millimètres à plusieurs mètres.
Cette faune a disparu en grande partie vers -544 Ma, n'ayant duré "que" 56 millions d'années.
Quittons maintenant l'ère du Précambrien et entrons dans la période du Cambrien (-541 à -485 Ma).
 |  |
| Reconstitution artistique de la faune d'Ediacara sur le fond marin. Photo Pierre-André Bourque d'une vitrine du Smithsonian Museum à Washington, 1995 | Reconstitution artistique de la faune des schistes de Burgess sur le fond marin. Un anomalocaris a capturé un trilobite. Merci à EnFOrE |
En 1886, un géologue travaillant à la construction d'une ligne de chemin de fer à travers le Canada remarqua la présence abondante de fossiles dans un endroit proche du mont Burgess dans les Montagnes Rocheuses canadiennes en Colombie-Britannique. Averti, un employé de la Geological Survey of Canada s'y rendit et vérifia la présence d'espèces inhabituelles telles qu'éponges, vers, et des sortes de crabes bizarres (qui plus tard se révéleront être une espèce inconnue, les anomalocarididés, nom signifiant étrange crevette). C'est ce qui décida le paléontologue américain Charles Doolittle Walcott (1850 - 1927) à explorer le site en 1909, où il trouva un ensemble impressionnant de fossiles. Ce site est classé au patrimoine mondial de l'UNESCO (Parcs des montagnes rocheuses canadiennes, 51°25'14" N, 116°30'20" O).
Walcott mit à jour plus de 65'000 exemplaires. Bien que situés à 2300 m d'altitude, ce sont des fossiles marins. En effet, le sol, fait de sédiments marins, a été soulevé au cours de la formation de montagnes. Il est daté de 515 Ma environ. Dans les années 1970, l'étude fut reprise par le paléontologue britannique Harry B. Whittington (1916 - 2010) et son équipe, qui s'affairèrent à décrire de multiples espèces, classées en 120 genres.
Leur diversité est bien plus grande qu'à Ediacara. La majorité des fossiles qu'on y trouve présente des morphologies bizarres. Parmi ceux-ci, une partie est rattachée à des embranchements connus à l'heure actuelle, possédant des squelettes externes (exosquelette) c'est-à-dire des coquilles ou des carapaces (éponges, algues, trilobites, mollusques, etc.). Une part plus importante d'entre eux sont des organismes à corps mou, sans squelette. Beaucoup ne ressemblent à aucune espèce actuelle et appartiennent à des groupes inconnus.
Whittington et ses collaborateurs sont arrivés à la conclusion importante que la faune du schiste de Burgess diffère fantastiquement de celle de la vie actuelle. Bien qu'elle comprenne beaucoup moins d'espèces, elle montre que les plans d'organisation anatomique présentent une diversité bien plus grande que les animaux actuels. Presque tous les embranchements modernes y font leur apparition.
Des fossiles semblables à la faune de Burgess ont été identifiés dans plusieurs autres gisements du monde entier: aux États-Unis, en Australie, en Chine. Celui de Sirius Passet au Groenland (82°47'36"N; 42°14'42"O), daté de -518 Ma, a été découvert en 1984. Cela montre que cette faune a occupé une grande partie des océans de la planète. Ces découvertes ont étendu la durée d'existence de cette faune sur tout le Cambrien, depuis environ 542 Ma jusqu'à 485 Ma.
L'un des dépôts les plus remarquables est celui des schistes de Maotianshan, en Chine, à Chengjiang (24°40'8"N, 102°58'38"E). Il a été découvert en 1984 par Hou Xian-guang. Ce site est inscrit au patrimoine de l'humanité. Il contient des fossiles d'espèces marines variées dont certaines sont communes avec les schistes de Burgess. Mais ils sont plus anciens, datant d'environ 525 Ma.
L'intérêt formidable du gisement chinois de Maotianshan, ainsi que des schistes de Burgess et de Sirius Passet au Groenland, est l'exceptionnel état de conservation des parties molles des animaux. On obtient de nombreux détails anatomiques d'invertébrés et de vertébrés. Parmi les nombreuses espèces, se trouvent une grande diversité d'algues, d'éponges, de cnidaires (sortes de méduses), de vers, d'arthropodes, des lamproies antiques appelées agnathes, et d'autres formes inconnues...
Pendant ce bref épisode de la vie sur la Terre, il y a eu un foisonnement intense de créations d'espèces et de plans d'organisation des organismes. C'est pourquoi cet épisode a reçu le qualificatif de grande explosion du Cambrien, ou même parfois big bang de la vie.
Les scientifiques ont remarqué que cette explosion correspond à une brusque augmentation du taux de l'oxygène dans l'atmosphère. C'est la deuxième, la première ayant eu lieu il y 2400 Ma (voir chapitre précédent).
 |  |
| Trilobite Kuanyangia pustulosa. Trouvé dans les schistes de Maotianshan. Merci à Dr Sam Gon III (A guide to the orders of trilobites) | Haikouella lanceolata, un vertébré trouvé dans les schistes de Maotianshan. Longueur environ 2 cm. Dessin de Stanton F. Fink (Apokryltaros). Merci à wikimedia |
Parmi les nouvelles espèces apparues dans les schistes de Maotianshan et de Burgess aux alentours de -570 Ma, les arthropodes figurent en bonne place, y compris le trilobite qui en est une sorte de vedette.
Le nom arthropode indique que les animaux ont des pattes faites de segments articulés (plus précisément, c'est le corps qui est constitué de plusieurs segments articulés). Chaque segment du corps est recouvert d'une carapace et muni d'une paire de pattes ou appendices articulés.
Actuellement, l'embranchement des arthropodes rassemble les araignées, les crustacés, les insectes, les scorpions, donc un nombre considérable représentant 80% des espèces vivantes. Alors qu'on dénombre 4 types d'organisation morphologique parmi les arthropodes actuels, on en a découvert 24 dans les schistes de Burgess, ce qui montre la variété existant à l'époque.
Les trilobites sont des arthropodes devenus populaires pour les chercheurs de fossiles car ils sont très répandus dans tous les continents sous des formes très diverses. On compte plus de 18'000 espèces échelonnées sur une longue période. On en trouve déjà quelques-uns dans les faunes d'Ediacara, de Maotianshan, de Burgess et de Sirius Passet. Leur taille varie entre 1 mm et 70 cm, avec une moyenne de 2 à 7 cm. Ils ont disparu il y a 250 Ma lors d'une extinction de masse à la fin du Permien (voir plus loin).
La fin du Cambrien est marquée par une réduction sensible du nombre d'espèces existantes.
Mais dans les 40 millions d'années qui suivent et qui constituent la période de l'Ordovicien (-485 à -444 Ma), la vie se développe et se diversifie de plus belle.
De nouveaux arthropodes font leur apparition. Certains sont bien plus grands que ceux de l'époque précédente, avec de fortes carapaces, tels les Euryptérides ou scorpions des mers dotés de fortes pinces.
Une équipe de chercheurs de l'Université de Yale a découvert en 2015 au fond d’un cratère de l'Iowa aux États-Unis plusieurs exemplaires très bien conservés d'un Euryptéride qu'ils ont nommé Pentecopterus. Il vivait il y a 467 Ma. Il est impressionnant avec ses 1,7 m de long.
Sources: On a retrouvé l'ancêtre des araignées, Slate, 1/09/2015; The oldest described eurypterid: a giant Middle Ordovician (Darriwilian) megalograptid from the Winneshiek Lagerstätte of Iowa, James C. Lamsdell, Derek E. G. Briggs, Huaibao P. Liu, Brian J. Witzke and Robert M. McKay, 2015, BMC Evolutionary Biology, 15, 169
La limule est un autre arthropode marin, remarquable parce que la famille est toujours représentée actuellement par 4 espèces (classe des xiphosures signifiant queue en forme d'épée).
La limule moderne mesure environ 30 cm de long et ressemble à un crabe. Son sang est de couleur bleue, due à la présence de cuivre. Il a la capacité de détecter des bactéries, ce qui a fait l'objet de recherches scientifiques. À cause de cela, leur pêche par les laboratoires nuit à l'espèce et la menace d'extinction.
C'est vraiment désolant, car la limule a été en mesure de traverser des centaines de millions d'années, y compris plusieurs extinctions massives, sans que leur morphologie soit modifiée. C'est pourquoi elle est taxée de fossile vivant. Cette longévité est attribuée au fait que sa constitution primitive lui procure de grandes facultés d'adaptation. Elle peut vivre dans des milieux divers, et résiste hors de l'eau.
On trouve de nombreux fossiles de limules au cours des différentes ères géologiques. Le plus ancien connu remonte à 445 millions d'années. Il a été découvert par une équipe de paléontologues canadiens, dans le Manitoba, province du Canada. Il mesure environ 4 cm.
Sources: Limulidae dans Wikipédia; La limule: un animal vieux de 450 millions d'années menacé d'extinction, Sciences et Avenir, 15/06/2016; The oldest horseshoe crab: A new xiphosurid from Late Ordovician Konservat-Lagerstätten deposits, Manitoba, Canada, 2008, Rudkin D. M., G. A. Young, and G. S. Nowlan. Palaeontology, 51, 1; Oldest Horseshoe Crab Fossil Found, 445 Million Years Old, Royal Ontario Museum, Science Daily, 8 février 2008
 |  |
| Limule vivante Tachypleus gigas échouée sur une plage indienne. Photographie de Shubham Chatterjee. Merci à Wikipédia | Un euryptide ou scorpion des mers, l'Acutiramus ou Pterygotus cummingsi ou buffaloensis. Dessin de Stanton F. Fink (Apokryltaros). Merci à wikimedia |
À la fin de l'Ordovicien, vers -444 Ma, environ 80% des espèces disparaissent, y compris une partie des trilobites. Les chercheurs pensent que cela est dû à une baisse de température importante consécutive à une glaciation.
Après cette réduction massive du nombre d'espèces, la diversité de la vie marine se déploie à nouveau au cours du Silurien (-444 à -419 Ma) et du Dévonien (-419 à -359 Ma)
On y retrouve les arthropodes en abondance. Parmi eux les euryptérides croissent en variétés. Le ptérygotus (ou acutiramus) pouvait dépasser 2 m de long.
Les ammonites, qui sont des mollusques de la classe des pieuvres et calmars, avec une coquille enroulée, apparaissent à la fin du Dévonien et deviennent abondantes après -250 Ma.
Les requins, poissons à squelette cartilagineux, apparaissent au Dévonien, il y a 420 Ma.
Vers -380 Ma, ils sont dominés par des poissons à quatre nageoires rigidifiées par un squelette osseux.
 |  |
| Fossile d'ammonite Perisphinctes Exposition du Muséum d'Histoire Naturelle de Berlin. Merci à Wikimedia, et I. Masur | Vue d'artiste du poisson vertébré Materpiscis. Taille environ 30 cm Par Sularko. Merci à Wikimedia |
Sources: Les premiers tétrapodes, Dinosoria; Ammonites, Wikipédia; Histoire évolutive des poissons cartilagineux, Wikipédia
Alors qu'auparavant tous les animaux vivaient dans la mer, des animaux terrestres font leur apparition. Ils profitent des plantes primitives qui s'installent sur les terres émergées devenues habitables. C'est ainsi que des acariens et des scorpions deviennent terrestres.
Une importante extinction massive a lieu au Dévonien, entre environ -380 et -360 millions d'années. Elle affecte plus de 70 % des espèces vivantes.
La cause de cette extinction reste inconnue: glaciation brève, impact de météores, éruptions volcaniques ou autre causes, rien ne permet de trancher.
Pendant les périodes qui suivent, le Carbonifère (-359 à -299 Ma) puis le Permien (-299 à -252 Ma), les océans sont peuplés de poissons, de requins, de coquillages. Les trilobites se font plus rares et finissent par disparaitre.
De grands arbres font leur apparition sur la terre émergée et des espèces terrestres y prolifèrent. On y voit les premiers amphibiens, les premiers reptiles, des mille-pattes, des araignées, et des insectes volants.
Vers -360 Ma, c'est l'époque de l'apparition des amphibiens. Un amphibien (également nommé batracien) présente des caractéristiques qui lui permettent de vivre dans l'eau et d'autres qui lui permettent de vivre dans l'air. Ils sont dotés de quatre membres et d'un cou. Un exemple est l'Ichthyostega, dont la respiration utilise à la fois des branchies comme les poissons, et des poumons.
En Allemagne, près de la ville d'Odernheim-am-Glan, les grès rouges de la strate Rotliegend (environ -300 à -270 Ma) sont riches en fossiles d'amphibiens. En 1847, le paléontologue allemand Georg August Goldfuss y a trouvé des Sclerocephalus qui pouvaient atteindre une longueur de 1,5 m.
 |  |
| Représentation artistique de l’amphibien Ichthyostega stensiosei par Nobu Tamura, 2007. Merci à Wikimédia | Fossile de l'amphibien Sclerocephalus haeuseri Photo de Günter Bechly. Musée national d'histoire naturelle de Stuttgart (Allemagne). Merci à Wikimédia |
Les premiers reptiles viennent tenir compagnie aux amphibiens. Le plus ancien reptile connu est Hylonomus lyelli. D'une longueur d'environ 20 cm, l'hylonomus a vécu il y a environ 315 Ma. Il a été découvert en 1852 par le géologue canadien sir William Dawson (1820 - 1899) dans les falaises fossilifères de Joggins en Nouvelle-Écosse, une province de la côte atlantique du Canada (site classé au patrimoine mondial de l'UNESCO).
Viennent ensuite de grands reptiles, ancêtres des dinosaures.
|
|  |
| Un reptile, l'Hylonomus Dessin de Ghedoghedo d'après une image du musée d'histoire naturelle de Stuttgart. Merci à Wikimédia | Un reptile, l'édaphosaure, pouvant mesurer jusqu'à 3 m de long. À ses pieds un amphibien le Platyhystrix. Les deux sont pourvus de voiles thermorégulatrices. Dessin de Dmitry Bogdanov, 2007. Merci à Wikimédia |
Les milles-pattes (myriapodes) prolifèrent. Arthropleura est le plus grand ayant jamais vécu, avec une longueur dépassant 2 m.
 |  |
| Arthropleura, mille-patte géant du carbonifère Source: Animal Picture Society | Fossile de l'araignée Planomartus krejcii, trouvé à Rakovnik, République tchèque. Longueur du corps environ 2 cm. Photo de Stanislav Stamberg, Paleontological Department du musée national de Prague, extraite de Carboniferous and Permian faunas and their occurence in the limnic basins of the Czech Republic, 2008 |
Dans la région de la Bohême en République Tchèque, de nombreux fossiles de différentes espèces datant du Carbonifère ont été découverts dans les sédiments de diverses localités comprenant des mines de charbon (ce qui est d'ailleurs à l'origine du nom carbonifère) et de minerai de cuivre.
D'abord un scorpion en 1935. Puis le paléontologue tchèque Antonin Fritsch (1832 - 1913) effectua une étude systématique qui a fourni les bases d'une recherche qui est poursuivie jusqu'à maintenant.
Dans ce bassin, on a trouvé des coquillages, des insectes, des amphibiens, des reptiles, des myriapodes et des araignées nommées arthromygales. D'autres trouvailles du même type ont été faites en Ukraine et en Russie, mais aussi dans d'autres terrains de l'Europe et de l'Amérique du Nord.
Les premiers insectes à 6 pattes, sans ailes, sont datés d'environ -395 Ma (Dévonien) mais ils ont probablement été présents avant, vers -415 Ma.
Des insectes avec ailes sont présents vers -385 Ma. Ils se développent au Carbonifère. à partir de -320 Ma. Les fossiles des calcaires d'Elmo au Kansas (États-Unis), découverts en 1899 et datés de -300 à -250 Ma, en ont fourni des milliers d'exemplaires en bon état.
Dans les mines de charbon de Commentry, dans l'Allier (France), l'entomologiste et paléontologue Charles Brongniart (1859 - 1899) a découvert en 1885, sur les indications des mineurs, l'empreinte d'une libellule géante dans les schistes de l'exploitation datés de -300 Ma. Nommée Meganeura, elle a une longueur de 30 cm et une envergure de 70 cm. Les schistes de Commentry offrent une large palette d'insectes aux ailes déployées.
En 2013, une équipe du CNRS a publié une étude sur des fossiles découverts dans une ancienne mine de charbon à Avion (Pas de Calais, France), dans une strate datée de -315 à -307 Ma. Ils y ont trouvé les plus anciens représentants connus des hémiptères (punaises et cigales), des coléoptères (scarabées, etc.) et des hyménoptères (guêpes et fourmis), groupes qui constituent une part très importante des espèces connues actuellement sur Terre. Leur taille est petite (longueur des ailes de 3 à 25 mm), ce qui montre qu'à cette période se côtoyaient des petits insectes et des très grands.
 |  |
| Fossile de Libellule Meganeura monyi Moulage d'un fossile original de Ch. Brongniart à Commentry déposé au Muséum d'Histoire Naturelle de Toulouse. Reproduit au Muséum des Sciences Naturelles de Bruxelles, Belgique. Merci à Wikimedia | Libellule Dunbaria fasciipennis, trouvée dans les calcaires d'Elmo (USA). Envergure environ 3 cm. Merci à Excursion zoologique aux alentours du Lac Baïkal |
Sources: The
Carboniferous: Arthropleura, dans Evolution of life; Revisional study of the order of paleodictyoptera in the upper carboniferous shales of Commentry, France, Jarmila Kukalova, Psyche: part I, 1969, 76, 2, 163, part
II, 1969, 76, 4, 439, part III, 1970, 77, 1, 1
Dans l’ombre des géants du Carbonifère: les plus anciens représentants des insectes modernes découverts dans un terril du nord de la France, CNRS Muséum national d'Histoire Naturelle, Communiqué de presse du 16 octobre 2013
The earliest-known holometabolous insects, Nel A., Roques P., Nel P. et coll., Nature, octobre 2013
Sclerocephalus dans le site Revolvy
Hylonomus dans le site du Musée Canadien de la Nature
On a Terrestrial Mollusk, a Chilognathous Myriapod, and some New Species of Reptiles, from the Coal-Formation of Nova Scotia, W. Dawson, 1860, Quarterly Journal of the Geological Society of London, 16, 268
Arachnids from the Carboniferous of Russia and Ukraine, and the Permian of Kazakhstan, Paul A. Selden, Dmitry E. Shcherbakov, Jason A. Dunlop, Kirill Yu. Eskov, 2014, Paläontologische Zeitschrift, 88, 3, 297
Carbonifère, dans le site Évolution biologique
Palaeodictyopteroida dans le site Palaeos
Wikipédia: Sclerocephalus; Hylonomus; Permien; Meganeura
Vers -250 Ma, date qui marque la fin de l'ère paléozoïque (ou primaire), se produit une nouvelle extinction de masse, la troisième grande extinction. Cette fois, elle prend une grande envergure: 75% des espèces terrestres et 95% des espèces marines connues disparaissent.
On ne sait pas ce qui a provoqué cette extinction, mais diverses causes sont envisagées: baisse du niveau de la mer, explosion volcanique, ou autre. Nous y reviendrons plus loin.
Le Mésozoïque est l'ère des dinosaures et des premiers mammifères.
Pendant la première période du Mésozoîque (le Trias, de −252 à −201 Ma), les reptiles marins peuplent les mers. Certains atteignent de grandes tailles. Les reptiles terrestres sont plus rares.
En outre, des reptiles volants (nommés ptérosaures) occupent le ciel dès -230 Ma.
C'est aussi l'époque des premiers rares dinosaures. Des fossiles d'Herrerasaurus ont été mis au jour en 1961 en Argentine dans la cordillère des Andes dans des strates datant de 230 Ma (formation d'Ischigualasto). L'animal a une longueur de 1 m environ. Toujours en Argentine dans la même formation, un fossile de dinosaure Eoraptor a été découvert en 1991. L'animal aurait entre 3 et 5 m de long.
 |  |
| Dinosaure Eoraptor. Découvert en 1991. Taille 1 m de long. Merci à Los
Dinosaurios más extremos | Dinosaure Herrerasaurus, découvert en 1961. Longueur 3 à 6 m. Merci à The Isle Wikia, d'après un modèle en 3D de Vlad Konstantinov |
Les premiers mammifères datent de la même époque, -230 Ma. Ils ressemblent à de petites souris. Les mammifères sont définis par des caractéristiques communes comprenant l'allaitement des petits, une peau couverte de poils, la respiration par des poumons. Citons adelobasileus, dont un fossile a été découvert au Texas en 1989.
Il y a 200 Ma, une nouvelle grande extinction, la quatrième, marque la fin du Trias. 50% de la faune disparait alors.
Dans la seconde partie du Mésozoïque (le Jurassique de -201 à -145 Ma et le Crétacé de -145 à -66 Ma), les espèces de dinosaures se multiplient et les reptiles volants sont nombreux dans le ciel.
 |  |
| Adelobasileus cromptoni, un des premiers mammifères. Dessin de Nobu Tamura. Merci à Wikimedia | Ptéranodon, reptile volant. Œuvre artistique d'Heinrich Harder (1858-1935). Illustration pour Die Wunder der Urwelt 1912. Merci à Wikimedia |
Le ptérodactyle est l'un des reptiles volants les plus connus, car il est le premier découvert en 1784, en Allemagne. Il vivait vers -140 Ma. Son envergure était de 70 cm.
Le ptéranodon en est un autre, bien plus grand que le ptérodactyle avec une envergure pouvant atteindre 7 à 9 m. Il a vécu plus tard, il y a environ 80 Ma.
De nombreuses sortes de dinosaures peuplaient la terre. Certains atteignent une taille gigantesque, d'où leur nom de dinosaure (=lézard géant). D'autres ont des tailles plus modestes, comparables à celle d'un humain, et d'autres ne dépassent pas la taille d'une poule.
Dans la province chinoise de Lioaning, les terrains de la formation de Jehol datés de 125 Ma environ ont livré de nombreux fossiles de dinosaures, d'oiseaux, de mammifères et d'autres espèces en parfait état.
Ainsi, le Mei Long, découvert en 2004, est un dinosaure de la taille d'un canard. Il se tient debout et se déplace sur les pieds de ses deux membres postérieurs. Les dinosaures qui possèdent cette caractéristique sont nommés des théropodes.
Dans la même formation chinoise, le squelette presque complet d'un dinosaure nommé Sinornithosaurus a été découvert en 1999. De la taille d'un aigle, il a la particularité d'être recouvert de plumes. Il n'est pas certain qu'il pouvait voler.
 |  |
| Sinornithosaurus, dinosaure à plumes découvert en Chine en 2001. Musée géologique de Chine. Merci à Wikimédia | Deux fossiles de l'oiseau Confuciusornis sanctus. Photo d'Aduard Solà, exposition à Barcelone. Merci à Wikimedia |
D'autres fossiles de dinosaures à plumes ont été découverts, ce qui montre que la présence de plumes était peut-être fréquente. Le Yutyrannus huali atteint la longueur de 9 m. Ses plumes ressemblent au duvet d'un poussin. Il ne volait probablement pas. (Le plus gros dinosaure à plume découvert en Chine, Le Figaro.fr, 05/04/2012)
Dans la formation chinoise, on trouve également des fossiles de véritables oiseaux. Le Confuciusornis est l'un des mieux connus car on en a trouvé plusieurs centaines de squelettes. Il a la taille d'un corbeau.
Pendant le Crétacé, les petits mammifères se développent et les insectes se diversifient.
On a aussi trouvé des puces. Des fossiles ont été mis au jour en Chine, les uns sur le site paléontologique de Daohugou en Mongolie intérieure datant de 165 Ma, les autres dans la province de Liaoning près de la Corée du Nord datant de 125 Ma. Plus grandes que les puces actuelles, elles mesurent de 8 à 20 mm. (Diverse transitional giant fleas from the Mesozoic era of China, Diying Huang, Michael S. Engel, Chenyang Cai, Hao Wu, André Nel, Nature, 2012, 483, 201)
 |  |
| Fossiles de puces de la période jurassique (165 Ma). Merci à Nature News | Scène du Jurassique sur une ile du nord de l'Allemagne. On y voit un dinosaure Europasaurus holgeri, des iguanodons de passage, deux Compsognathus en arrière-plan et un Archaeopteryx à droite. Tableau de Gerhard Boeggemann. Merci à Wikimedia |
L'ère mésozoïque se termine en -66 Ma par une cinquième extinction massive, marquée par la disparition d'une majorité de dinosaures et de nombreuses autres formes de vie, y compris les ammonites et les reptiles marins. Plus de la moitié des espèces vivantes disparaissent.
Après la grande extinction de -66 Ma, commence l'ère Cénozoïque (du grec Vie récente), qui dure jusqu'à aujourd'hui. Elle regroupe le Tertiaire (-66 à -2,5 Ma) et le Quaternaire.
Elle est marquée essentiellement par la diversification des oiseaux, des insectes et des mammifères, qui prennent des formes plus complexes, ainsi que par le développement des primates et l'avènement de l'homme.
À titre d'exemples, je rapporte le récit de l'évolution du cheval, puis dans la section suivante, des hominidés et de l'homme.
Le cheval actuel appartient à la classe des mammifères et à la famille des Équidés, qui compte aussi les ânes et les zèbres. Les pattes des équidés se caractérisent par des segments très allongés. La main et le pied ne possèdent qu'un seul doigt.

Sifrhippus reconstitué. Musée d'histoire naturelle suédois. Merci à Wikimédia
Le représentant le plus ancien de cette famille a été découvert en Amérique du Nord, dans le bassin de la Bighorn dans le Wyoming. C'est le Sifrhippus sandrae (anciennement appelé Hyracotherium sandrae). Il a la taille d'un chat domestique.
Source: Quand l'ancêtre du cheval était petit comme un chat, Tristan Vey, Le Figaro.fr, 23/02/2012
Les spécialistes se disputent pour classer une autre espèce de cette époque, l'Hyracotherium, dans le même genre Sifrhippus ou non.
De la taille d'un petit chien, il a vécu en Asie, en Europe et en Amérique du Nord entre environ -53 et - 45 Ma. Ses pattes avant possèdent 4 doigts dont l'extrémité repose sur le sol et ses pattes arrière possèdent 3 doigts., et non un seul comme les équidés modernes.
Dès qu'un animal est classé dans une famille, les paléontologues recherchent son ancêtre commun à d'autres familles. Aussi déploient-ils de grands efforts pour classer leurs trouvailles.
Cela vient du fait qu'ils se positionnent systématiquement dans le cadre de la théorie de l'évolution néo-darwiniste, basée sur l'idée de transformation génétique des espèces à partir d'un ancêtre. Mais cette théorie est purement spéculative, comme je l'expose dans le chapitre suivant.
En conséquence, sont nées des controverses sur l'appartenance des deux espèces que je viens de présenter. Pour les uns, c'est l'Hyracotherium leporinum qui est l'ancêtre, et pour les autres, il appartient à la famille des paléothères et c'est Sifrhippus sandrae qui est l'ancêtre.
Classer un animal dans une famille plutôt qu'une autre dépend des traits communs que l'on choisit pour définir la famille, et c'est subjectif (voir en annexe La classification des espèces résulte de choix subjectifs).
En Amérique du Nord, leur succèdent différents genres dont Eohippus, Orohippus, Mesohippus, Meryhippus, Miohippus.
Le Mesohippus a vécu il y a environ 30 à 40 Ma. Il ressemble à un petit cheval, avec des pattes avant à 3 doigts. Le Merychippus a 80 cm de haut. Ses pattes ont 3 doigts, mais le doigt central devient plus important. Il a vécu il y a 20 à 17 Ma. L'Equus, le cheval actuel, apparait vers -4 Ma. Il a un seul doigt, le sabot.
Différents types de chevaux ont migré vers l'Europe et l'Asie. Ceux de l'Amérique du Nord sont tous éteints. Ceux qui y vivent actuellement ont été importés par les Espagnols il y a 500 ans.
Les primates sont des mammifères incluant les singes et les humains, mais aussi les petits animaux surprenants que sont les tarsiers.
Les caractères communs qui les définissent, en plus de ceux des mammifères, sont des yeux qui regardent vers l'avant, des membres à 5 doigts terminés par un ongle plat, des mains avec un pouce qui leur permettent de prendre des objets avec facilité.
Le plus ancien primate connu avec certitude est un squelette entier de Archicebus achilles, découvert en Chine en 2003 par le paléontologue chinois Xijun Ni dans la province de Hubei. Il est daté de 55 Ma. Il mesure environ 7 cm pour le corps sans la queue.
Il possède des caractéristiques qui l'apparentent à la fois aux tarsiers par ses proportions générales et ses dents, et aux anthropoïdes (singes) par ses pattes arrières. Si les paléontologues n'avaient trouvé que les dents, ils l'auraient classé comme tarsier, raconte Paul Tafforeau, co-auteur de l'étude. De quoi faire réfléchir sur la relativité d'une classification (voir en annexe La classification des espèces résulte de choix subjectifs).
Sources: Découverte en Chine du plus proche ancêtre des primates, Sciences et Avenir, 07/06/2013; Découverte du plus vieux primate du monde, Le Figaro.fr, 07/06/2013; The oldest known primate skeleton and early haplorhine evolution, Xijun Ni, Daniel L. Gebo, Marian Dagosto, Jin Meng, Paul Tafforeau, John J. Flynn & K. Christopher Beard, Nature, 2013, 498, 60
De nos jours, différentes espèces de tarsiers peuplent les forêts des iles d'Asie du sud-est (Philippines, Sumatra, Bornéo, Sulawesi). Ils sont étonnants par leurs grands yeux qui emplissent une grande partie de leur face. Petits, ils mesurent environ 23 cm pour le corps sans la queue. Leurs grands pieds (les os du tarse) leur permettent de bondir d'arbre en arbre à la manière d'une grenouille pour atterrir pile sur leur proie, qui consiste en petits insectes comme des sauterelles. Malheureusement, ces espèces sont menacées par l'afflux des touristes et une fondation a été créée pour leur sauvegarde.
Des fossiles de tarsiers ont été découverts en Asie, en Europe et en Amérique du Nord. L'espèce la plus ancienne, Tarsius eocaenus, a été découverte en 1992 dans une terrain fossilifère daté de 45 Ma, dans la province de Jiangsu en Chine, en compagnie d'autres primates. De l'animal, il ne reste que des dents. Le fossile atteste que la formule dentaire est restée inchangée pendant 45 Ma. (A Diverse New Primate Fauna from Middle Eocene Fissure-Fillings in Southeastern China, K. Christopher Beard et al., Nature, 1994, 368, 6472, 604)
 |  |
| Un tarsier actuel. Réserve Philippine Tarsier Sanctuary, province de Bohol, Philippines. Merci à Carlo
Lorenzo | Reconstitution artistique du premier primate connu, Archicebus achilles. Photo de Xijun Ni/AP |
Les singes sont appelés anthropoïdes parce qu'ils ressemblent aux êtres humains (grec: anthropos). Ce groupe de primates comprend de nombreuses espèces très diversifiées. On distingue d'une part les petits singes dotés d'une queue, et d'autre part les grands singes sans queue (singes anthropomorphes ou hominoïdes - ne pas confondre avec humanoïdes).
Les fossiles des plus anciens singes connus ont été trouvés en Chine en 1995, dans le basin Yuanqu, province Shanxi, par des paléontologues états-uniens et chinois sous la direction de K. Christopher Beard. Il s'agit de la dentition d'un très petit primate baptisé Eosimias centennicus. Il est daté de 45 Ma. (Earliest complete dentition of an anthropoid primate from the late Middle Eocene of Shanxi Province, K.C. Beard et al., Science, 1996, 272, 82)
Des espèces anciennes similaires ont été découverts dans plusieurs lieux d'Asie, dont la Thaïlande et le Myanmar (Birmanie). Citons en particulier, des fossiles du petit singe Bahinia pondaugensis, datés de 40 Ma en Myanmar, découverts en 1999 par le paléontologue Jean-Jacques Jaeger et son équipe.

Squelette du singe Proconsul reconstitué. Musée d'anthropologie, Université de Zürich, Suisse. Merci à Wikimedia (N. Guérin)
On trouve des fossiles de petits singes très semblables en Afrique (Égypte, Algérie, Lybie), les plus anciens datant de 40 Ma.
Ainsi le petit singe Aegyptopithecus zeuxis, datée de -30 Ma, a été trouvé en 1966 dans l'Oasis du Fayoum en Égypte, par le paléontologue américain Elwyn Simons (1930 - 2016).
L'espèce Afrotarsius libycus, datant de 39 Ma, a été découverte en Libye en 2010 sur le site de Dur At‐Talah par une équipe dirigée par le paléontologue français Michel Brunet (né en 1940).
Les grands singes hominoïdes font leur apparition vers -26 Ma. Les fossiles de l'espèce Proconsul, exhumés au Kenya et en Ouganda, datent de 20 Ma environ. Le principal fossile de Proconsul, vieux de plus de 18 Ma, a été découvert en 1948 par la paléontologue Mary Leakey (1913 - 1996) et son équipe sur l'ile de Rusinga, dans le lac Victoria au Kenya.
Michel Brunet et son équipe ont aussi mis au jour en 2001, dans le désert du Djourab au Tchad, le crane fossilisé d'un singe daté de 7 Ma selon la méthode de datation au béryllium 10 radioactif.
Baptisé Sahelanthropus tchadensis, surnommé Toumaï, son analyse suggère qu'il s'agit d'un grand singe bipède, qui pouvait grimper aux arbres. Alors que les autres primates qui se déplacent en position debout le font seulement de façon occasionnelle, ces singes se déplacent debout sur leurs deux pieds de façon permanente.
Les grands singes bipèdes, les australopithèques décrits ensuite et les espèces du genre Homo forment la famille des hominidés. Toumaï serait donc le plus ancien hominidé connu.
Un autre, Orrorin tugenensis, découvert au Kenya en 2000 par la paléontologue française Brigitte Senut (née en 1954), gisait dans des sédiments de 6 Ma.
Ces deux découvertes ont fait reculer d'un grand bond la date d'apparition des hominidés, auparavant estimée à 4 Ma. De telles révisions sont fréquentes en paléontologie. Elles devraient conduire les scientifiques à une certaine circonspection avant d'affirmer que telle espèce est apparue à telle date. Il est en effet toujours possible que des découvertes ultérieures viennent changer la donne. On peut seulement affirmer connaitre le plus ancien fossile connu.
En 1924, Raymond Dart (1893 - 1988), anthropologue d'Afrique du Sud, identifia la découverte à Taung près de Kimberley en Afrique du Sud du crâne d'un jeune hominidé. Baptisé l'enfant de Taung (Australopithecus africanus), c'est le premier exemplaire du genre Australopithèque (signifiant "singe du sud"), daté de 4 Ma. Le site est maintenant classé comme patrimoine mondial de l'UNESCO (Hominidés fossiles d’Afrique du Sud).
Par la suite, on a trouvé beaucoup d'autres exemplaires d'australopithèques dans plusieurs pays d'Afrique, appartenant à plusieurs espèces. Le plus célèbre est Lucy (Australopithecus afarensis), daté de 3,2 Ma, mis au jour en Éthiopie par le géologue français Maurice Taïeb avec Yves Coppens et Donald Johanson, en 1974. Les australopithèques ont disparu il y a environ 1 Ma.
En 1856, les ouvriers d'une carrière trouvent, dans une grotte de la vallée de Neander en Allemagne, des ossements et le crâne d'un homme préhistorique. C'est l'homme de Néandertal Homo neanderthalensis.
Les plus anciens exemplaires connus de néandertaliens, exhumés en Espagne dans le site d'Atapuerca, datent de 430'000 ans. D'autres auraient vécu en Europe, au Moyen-Orient et en Asie Centrale jusqu'à -35'000 ans. Son cerveau est volumineux avec un volume de 1450 cm3. Il fabrique des outils (pointes, racloirs).
Mais on s'apercevra vite que le genre Homo a existé bien avant l'homme de Néandertal avec deux espèces: Homo Habilis (=homme habile) et Homo Rudolfensis, qui ont vécu en Afrique orientale et en Afrique du Sud entre -2,5 et -1,5 Ma. Homo habilis fabriquait des outils en galets taillés.
Par rapport aux australopithèques, le genre Homo est caractérisé par un crâne plus volumineux (supérieur à 550 cm3), une denture plus étroite, et quelques autres détails anatomiques qui lui donne une silhouette plus gracile.
Différentes espèces d'Homo ont été identifiées pour les périodes échelonnées depuis 2 Ma jusqu'à -50'000 ans.
Homo ergaster a vécu en Afrique entre -1,9 et -1 Ma.
Homo erectus a vécu en Eurasie entre -1,2 Ma et -300'000 ans. Un exemple de fossile d'Homo erectus est l'homme de Tautavel, un crâne daté de 320'00 ans trouvé en 1971 dans une grotte près de Tautavel (Pyrénées orientales).
Homo rhodesiensis a vécu en Zambie (Afrique) entre -700'000 et -300'000 ans.
Homo georgicus est représenté par plusieurs crânes découvert en 2005 en Géorgie sur le site de Dmanisi. Ils sont datés de -1,8 Ma. Or bien que trouvés au même endroit et faisant probablement partie du même groupe humain, ces crânes ont des particularités anatomiques différentes.
Aussi, selon l'étude d'une équipe internationale de chercheurs menée par le Muséum National de Géorgie à Tbilisi en 2013 sur l'un des crânes, les distinctions entre les espèces anciennes du genre Homo qui vivaient il y a environ 2 millions d'années (Homo rudolfensis, Homo habilis, Homo ergaster, Homo georgicus, etc.) n'ont pas de sens, car ces hominidés font tous partie de la même espèce, Homo erectus. En effet, le crâne étudié présente une boîte crânienne relativement petite (546 cm3) comme Homo habilis, une face allongée comme Homo erectus, et de grandes dents comme Homo rudolfensis. (Un crâne préhistorique bouleverse l'évolution humaine, Science et Avenir, 18/10/2013)
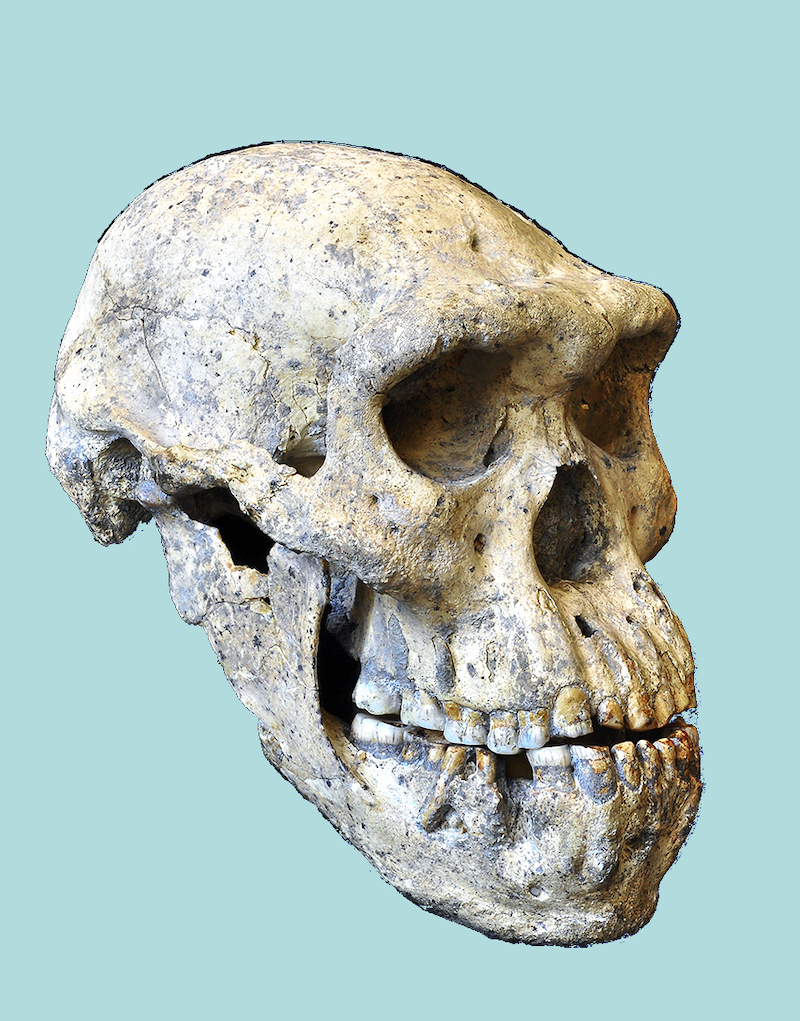 | 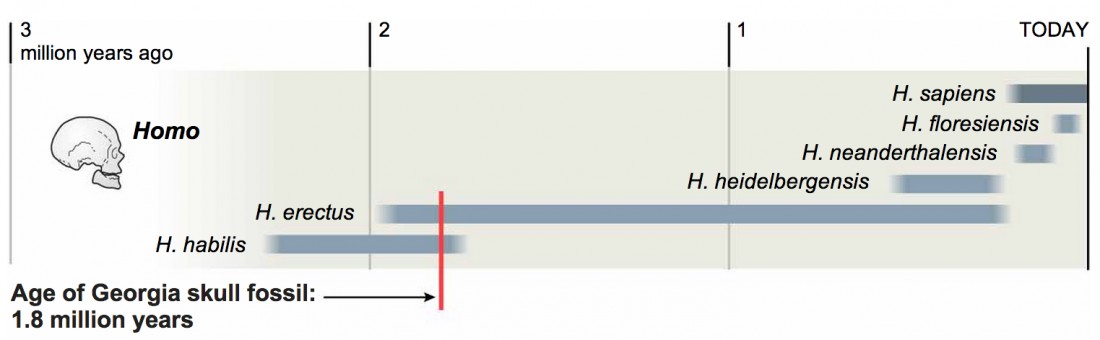 |
| Crâne découvert à Dmanisi. Photo G. Bumbiashvili, Georgian National Museum. Merci à Sciences et Avenir | Nouvel arbre généalogique des hominidés proposé par les chercheurs du Muséum National de Géorgie. Crédit NY Times. Merci à Sciences et Avenir |
Homo sapiens est le dernier Homo apparu. C'est nous. C'est la seule espèce d'homo encore existante.
Les fossiles d'homo sapiens les plus anciens connus ont été trouvés à Djebel Irhoud, au Maroc, par l'équipe du paléoanthropologue français Jean-Jacques Hublin (né en 1953) avec Abdelouahed ben-Ncer entre 2004 et 2016. Ils les ont datés de 300'000 ans. (New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens, J.J. Hublin et al., Nature, 2017, 546, 289)
Cela a reculé d'un coup l'apparition d'Homo sapiens de 100'000 ans, car précédemment les plus anciens fossiles étaient deux crânes datés de 195'000 ans trouvés en Éthiopie en 1967.
En Europe, on le trouve à partir de 45'000 ans. L'un des plus célèbres est l'homme de Cro-Magnon (-35'000 ans), du nom du lieu-dit où il a été trouvé en 1868 en Dordogne, aux Eyzies-de-Tayac dans un abri sous roche.
Sources: L’homme descend-il d'un primate passé d'Asie en Afrique? François Savatier, Pour La Science, 10/11/2010; Les ancêtres de la lignée humaine étaient-ils Asiatiques? Quentin Mauguit, Futura, 06/06/2012; De l’origine des anthropoïdes à l’émergence de la famille humaine, M. Brunet et J.J. Jaeger, C.R. Palevol, 2017, 16, 2, 189; Les derniers préhumains et les premiers humains, Yves Coppens, C.R. Palevol, 2017, 16, 2, 196; Les Anthropoïdes Éocènes, Jean Jacques Jaeger, CNRS
Avec le recul, nous pouvons embrasser la succession des entrées en scène de la vie animale depuis la bactérie jusqu'à l'être humain. Il ne nous échappe pas que des formes de vie de plus en plus complexes se sont manifestées sur cette Terre. Est-ce le signe d'une finalité?
La théorie de l'évolution darwiniste a conditionné les esprits à penser que les espèces se transforment progressivement vers une meilleure adaptation à l'environnement en développant de nouvelles capacités afin de survivre plus facilement. Les espèces moins adaptées feraient place aux nouvelles espèces mieux adaptées (voir les détails dans le chapitre suivant).
La complexité croissante serait-elle le témoignage d'une meilleure adaptation? Les formes complexes seraient-elles supérieures aux formes simples? Rien ne permet de l'affirmer, bien au contraire.
D'une part, les apparitions de nouvelles espèces ne sont pas aussi progressives que la présentation de leurs apparitions le laisse supposer - y compris celle de cet article.
Une espèce simple peut apparaitre postérieurement à une espèce complexe. En fait il y a des apparitions très variées, qui ne suivent pas un ordre linéaire.
Ainsi, on ne sait pas vraiment si les procaryotes (sans noyau) ont précédé les eucaryotes (avec noyau), car on ne connait pas leurs dates d'apparition respectives. En comparant les séquences moléculaires d'ARN (voir article ADN moléculaire), des généticiens ont estimé que les archées (bactéries anciennes) et les eucaryotes sont bien plus anciens que les plus vieux fossiles bactériens, et qu'elles se sont développées parallèlement (selon Carl Woese, professeur de microbiologie, découvreur des archées, dans When did eukaryotic cells first evolve? What do we know about how they evolved from earlier life-forms? Scientific American).
Selon la théorie de l'endosymbiolose, développée par la biologiste états-unienne Lynn Margulis dans les années 1960, une cellule procaryote a absorbé et intégré une autre cellule procaryote (ceci parce qu'on place l'intention dans la cellule qui absorbe. Si on place l'intention dans la cellule qui s'incruste, alors, elle colonise). Au lieu de la digérer, elle a conclu une association bénéfique pour les deux et la cellule absorbée est devenue un organite interne, mitochondrie ou chloroplaste.
Cette théorie est séduisante à cause de la similitude des organites dans la cellule eucaryote et des cellules procaryotes isolées. Mais rien ne prouve qu'elle soit vraie, dans la mesure où des théories alternatives tout aussi séduisantes ont été proposées.
Une théorie alternative a été suggérée en 2014 par David Baum, professeur de botanique aux États-Unis. Il affirme qu'une cellule procaryote aurait construit un corps autour d'elle, en devenant le noyau de la nouvelle cellule. (An inside-out origin for the eukaryotic cell, David A Baum and Buzz Baum, BMC Biology, 2014)
Mais les scientifiques sont tellement enfermés dans le cadre limité du néo-darwinisme, qu'ils supposent a priori que l'évolution va systématiquement dans le sens d'une plus grande complexité et que les organismes simples précèdent forcément les organismes complexes. Ils raisonnent en termes de lutte, alors qu'on peut penser en termes de coopération et de synergie.
Ils ont donc imaginé un mécanisme, largement accepté par la communauté, qui expliquerait le passage de la cellule procaryote à la cellule eucaryote: c'est l'endosymbiose, c'est-à-dire l'absorption d'une bactérie par une autre bactérie (voir l'encadré ci-contre).
D'autre part, rien ne permet de conclure qu'une structure complexe est mieux adaptée qu'une structure simple et va la remplacer. La structure eucaryote est-elle meilleure que la structure procaryote?
Même simple, la bactérie est un organisme hautement évolué par lui-même, incluant tous les mécanismes biologiques qui lui permettent de remplir des fonctions très diverses dans des environnements très variés. Certaines bactéries vivent dans des conditions extrêmes, invivables pour des eucaryotes (voir chapitre précédent, L'apparition de la vie).
Cela signifie que chaque espèce a ses propres capacités, et qu'elle se développe selon sa propre nature.
Enfin, l'apparition d'une espèce complexe n'élimine pas les espèces plus simples. Elles vivent ensemble en équilibre écologique.
La présence d'organismes eucaryotes dès 2400 Ma n'a pas éliminé les bactéries. Les deux ont vécu et vivent encore dans le même milieu.
Bien plus, ils sont associés et indispensables les uns aux autres. Ils vivent en synergie. Les bactéries sont indispensables à la vie des sols, sans lesquels les plantes ne survivraient pas. Il en est de même pour les animaux, y compris l'Être humain, qui vivent en symbiose avec des micro-organismes.
Le Pr Didier Raoult, microbiologiste français réputé, auteur de Dépasser Darwin (2010), affirme: On découvre que l'homme est un écosystème à lui tout seul, un monde dans lequel cohabitent des millions de micro-organismes. Cet écosystème ambulant évolue dans d'autres écosystèmes qu'il modifie et qui le modifient. (interviewé dans Le Point, Et si Darwin s'était trompé, 12/12/2011)
Autrement dit, les équilibres écologiques deviennent de plus en plus complexes, plus riches, en incluant des organismes qui font preuve d'inventivité très variée. Mais cela ne rend pas les autres inutiles. Chaque espèce a des capacités et des fonctions différentes et complémentaires dans le vaste écosystème.
L'étude des fossiles montre que le développement de la vie n'a jamais été uniforme et linéaire. Nous avons noté plusieurs fois des réductions importantes du nombre d'espèces, suivies de l'apparition soudaine d'abondantes espèces nouvelles.
Pour préciser cela, les paléontologues états-uniens J. John Sepkoski (1948 - 1999) et David M. Raup (1933 - 2015) ont établi en 1982 un catalogue précis de 3300 genres de fossiles marins vertébrés et invertébrés depuis 600 millions d'années et de leur variations en nombre en fonction de leur couche stratigraphique (Mass extinctions in the marine fossil record, D.M. Raup et J. JJ.Jr. Sepkoski, Science, 1982, 215, 4539, 1501).
Un catalogue plus complet de Sepkoski a été publié en 2002 (A compendium of fossil marine animal genera. Bull Amer Paleontol, 2002, 363, 560). Il comprend 37'000 genres d'animaux marins. Il est disponible en ligne comme base de données à l'université de Wisconsin-Madison.
N'y sont évidemment répertoriés que les fossiles qui ont été préservés et que des paléontologues ont eu la chance de trouver. Il y a donc lieu d'être prudent pour leur accorder une représentativité. Mais nous pouvons admettre qu'ils nous donnent une indication générale très intéressante.
À partir de ce catalogue, on a pu analyser les phases d'extinction massive et les variations de la biodiversité.
On parle d'extinction lorsque le nombre d'espèces a diminué de plus de 70% environ. On parle d'extinction massive lorsqu'elle a lieu sur la planète entière.
J'ai déjà mentionné 5 crises majeures dans le fil de cet article. Ces extinctions marquent des changements spectaculaires dans la vie animale de la planète, au point que les paléontologues ont retenu ces périodes comme bornes de leur division en ères géologiques.
Rappelons-les:
1. 445 Ma, fin de l'Ordovicien. À la limite entre l'Ordovicien et le Silurien, disparition de 27% des familles et 57% des genres d'animaux marins, ou 85% au niveau des espèces.
2. 370 Ma, fin du Dévonien. L'extinction du Dévonien élimine 19% des familles et 35 à 50% des genres d'animaux marins, ou 75% au niveau des espèces.
3. 250 Ma, fin du Permien. L'extinction du Permien-Trias est la plus massive. Près de 95 % de la vie marine disparait ainsi que 70 % des espèces terrestres (plantes, animaux).
4. 200 Ma, fin du Trias. L'extinction du Trias-Jurassique marque la disparition de 75 % des espèces marines, et 35 % des familles d’animaux.
5. 66 Ma, fin du Crétacé. L'extinction Crétacé-Tertiaire réduit les espèces de 50%.
À partir de leur catalogue de 1982, Raup et Sepkoski ont analysé la dynamique de l'évolution.
Outre les crises, des espèces apparaissent et d'autres disparaissent de façon continue. Le taux d'extinction continue est de l'ordre de 2 à 5 familles d'animaux marins par million d'années depuis 540 Ma.
Raup et Sepkoski ont attiré l'attention sur l'aspect périodique des extinctions. Ils ont pris en compte non seulement les extinctions massives mais un total de 12 extinctions depuis 250 Ma. Ils estiment la durée de ces cycles à 26 Ma. Les extinctions sont suivies d'un renouvellement de la faune et de la flore, caractérisé par une grande diversité écologique et morphologique. (Periodicity of extinctions in the geologic past, D.M. Raup and J.J. Sepkoski Jr, Proc Natl Acad Sci U S A, 1984, 81, 3, 801)
Les causes des extinctions devraient donc elles-mêmes être cycliques. Nous les explorerons plus loin.
Par la suite, plusieurs équipes ont analysé les mêmes données du catalogue, avec des méthodes statistiques différentes. A. Melott et R. Bambach en 2014, puis M. Rampino et K. Caldeira en 2015, ont confirmé la valeur de 27 Ma. D'autres ont trouvé des durées de cycle différentes, par exemple 35 Ma.
On peut tirer parti du catalogue pour dénombrer les espèces vivant à une époque précise: c'est la diversité biologique.
Courbe montrant le nombre de genres répertoriés dans les fossiles depuis 542 Ma. Le cycle de 62 Ma de la diversité des fossiles y est apparent. Merci à R. Muller. Voir aussi Wikimedia
Dans une Letter to Nature, les physiciens états-uniens Robert A. Rohde et Richard A. Muller (né en 1944) reprennent les données du catalogue de Sepkoski de 2002, et repèrent l'apparition et la disparition d'un genre. Ils évaluent ainsi la diversité biologique en fonction du temps, défini selon l'échelle de la Commission Internationale de Stratigraphie de 2004 (Cycles in fossil diversity, Rohde et Muller, Nature, 2005, 434, 208).
En soustrayant l'évolution continue lente, ils ont montré qu'une évolution oscillante lui était superposée. C'est un cycle de 62±3 Ma. Les 5 extinctions de Sepkoski se calent dans la phase descendante de ce cycle de 62 Ma.
Un autre cycle de 140±15 Ma lui est superposé.
Une analyse ultérieure par Bruce S. Lieberman et Adrian L. Melott a confirmé le cycle de 62 Ma en précisant qu'il est le même pour la biodiversité, pour l'apparition et pour l'extinction des genres (Considering the Case for Biodiversity Cycles: Re-Examining the Evidence for Periodicity in the Fossil Record, B.S. Lieberman, A.L. Melott, PLoS ONE, 2007, 2, 8, e759).
Les causes possibles de ces extinctions massives ont suscité de nombreuses interrogations. Et cela d'autant plus qu'il faut expliquer leur aspect périodique. Différentes réponses ont été suggérées, sans qu'aucune ne puisse être validée définitivement. Passons-les rapidement en revue.
Une grande glaciation provoque le retrait de la mer et entraine des modifications écologiques au-delà des capacités d'adaptation des espèces. Du gaz carbonique ou du sulfure d'hydrogène toxique est libéré des océans, et la teneur en oxygène baisse.
Éruption d'un super volcan en Sibérie.
Selon une étude de 2014 menée conjointement par le MIT et l'académie chinoise des sciences de Nanjing, une archée pathogène se serait développée de façon spectaculaire au Permien. (L’extinction de masse du Permien est-elle due à des micro-organismes ?, Futura-Science, 3 avril 2014)
En glissant sur la surface terrestre, les continents se rapprochent, les plateaux continentaux disparaissent au moment de la collision. Les courants océaniques sont modifiés.
Une équipe de chercheurs a entrepris des analyses de strontium dans les sols marins. Le rapport de la quantité des isotopes Sr87/Sr86 présente une périodicité de 59 Ma, proche des 62 Ma de la diversité des genres. Ils proposent que la Terre subit des pulsations régulières de son manteau ou des plaques tectoniques, qui pourraient se soulever comme un coeur battant. Elles affecteraient la chimie des océans.
Autre cause sérieusement envisagée: l'impact d'un ou de plusieurs météorites. Ils peuvent revenir cycliquement.
Depuis 1979, plusieurs équipes de chercheurs ont rapporté que le nombre de cratères formés par l'impact de comètes sur la Terre varie de façon périodique, avec une périodicité d'environ 36 à 38 Ma. (The period dichotomy in terrestrial impact crater ages, Richard B. Stothers, Mon. Not. R. Astron. Soc., 2006, 365, 178)
Rampino et Caldeira trouvent 26 Ma, ce qui correspond au cycle des extinctions de Raup et Sepkoski. (Periodic impact cratering and extinction events over the last 260 million years, Michael R. Rampino, Ken Caldeira, Monthly Notices of the Royal Astronomical Society, 2015, 454, 4, 3480)
Certains relient la périodicité des extinctions au mouvement du système solaire qui oscille verticalement par rapport au disque galactique. La demi-période d'oscillation est de l'ordre de 32 Ma environ (voir mon article Spirale, mouvement de vie). Cela perturberait régulièrement le nuage d'Oort et ses nombreuses comètes.
Le nuage d'Oort est une accumulation de comètes aux confins du système solaire, source de beaucoup de comètes qui le traversent. Il est situé beaucoup plus loin que la ceinture de Kuiper, autre source de comètes.
 |  |
| Représentation de la ceinture de Kuiper et du nuage d'Oort par rapport au système solaire. Merci au Herschel Space Observatory | Représentation de la voie lactée et de la trajectoire ondulante du soleil. Diagramme de C. Carreau-ESA. Merci à Nature |
Pour Napier et Wickramasinghe, lorsque le système solaire et son nuage d'Oort passent dans le disque galactique, ils rencontrent une densité stellaire et interstellaire plus grande, et cela provoque des flux de comètes. (Impact cratering and the Oort Cloud, J.T. Wickramasinghe, W.N. Napier, Monthly Notices of the Royal Astronomical Society, 2008, 387, 153)
Afin d'expliquer des signaux de rayons gamma en provenance du centre de la galaxie, détectés par le télescope Fermi Gamma-ray Space Telescope, les physiciens états-uniens Lisa Randall et Matthew Reece ont proposé qu'un disque de matière noire existe dans le plan galactique.
La matière noire est une matière hypothétique qui remplit l'univers. Elle est invisible, mais son existence pourrait rendre compte de phénomènes de gravité observables (voir mon article L'éther et l'énergie de l'espace selon la physique quantique).
Selon ces deux physiciens, c'est en traversant ce disque noir tous les 35 Ma que le soleil, dans ses oscillations verticales, provoque l'afflux de comètes vers la Terre, en perturbant le nuage d'Oort. (Dark Matter as a Trigger for Periodic Comet Impacts, Lisa Randall and Matthew Reece, arXiv 1403.0576v, 3 mars 2014)Medvedev et Mellot pensent que la traversée du plan galactique par le soleil engendre un flux de rayons cosmiques. (Do extragalactic cosmic rays induce cycles in fossil diversity? MV Medvedev et AL Melott, Astrophys J, 2007, 664, 879)
Les observatoires détectent fréquemment des bouffées brèves de rayons γ (ou sursauts gamma), extrêmement puissantes. Elles durent de quelques millisecondes à quelques minutes. Les plus intenses délivrent autant d'énergie que le soleil en 10 milliards d'années.
On détecte en moyenne un sursaut γ par jour (ou GRB - gamma ray burst). Leur cause reste débattue. Les scientifiques pensent qu'ils sont provoqués par l'explosion d'étoiles géantes comme les super-novas, ou par la collision d'étoiles à neutrons.
Le plus puissant sursaut a été enregistré le 16 septembre 2008 (Un sursaut gamma pulvérise le record des explosions cosmiques, L. Sacco, 23/02/2009). Un sursaut gamma en provenance de notre galaxie semble s'être produit en 774 (Un sursaut gamma aurait épargné la Terre au huitième siècle, L. Sacco, 23/01/2013).
Ces rayons énergiques, lorsqu'ils ne sont pas affaiblis par la distance, détruisent la couche d'ozone de la Terre et expose les organismes à l'action néfaste des rayons UV. De tels sursauts gamma pourraient avoir été responsables de destructions sur la Terre et avoir provoqué des extinctions massives. L'astrophysicien états-unien Brian Thomas suggère que c'est le cas de l'extinction de l'Ordovicien il y a 445 Ma.
Des explosions encore plus puissantes se produisent souvent dans le centre des galaxies. Depuis les années 1960, les astronomes ont observé que le noyau de certaines galaxies (galaxies de Seyfert et quasars) explosait en émettant des rayonnements intenses s'étendant sur tout le spectre électromagnétique: radio, lumineux, rayons X et γ. Dans certaines galaxies, les émissions observées sont équivalentes à des milliards d'explosions de super-nova.

Dr Paul A. LaViolette
Merci à Starburst Foundation
Le physicien états-unien Paul A. LaViolette (né en 1947), président de Starburst Foundation, est connu pour ses recherches sur une nouvelle théorie de la matière qu’il appelle la Cinétique subquantique. Dans son mémoire de doctorat d'astrophysique en 1983 et dans son livre Earth under Fire, il a montré que de telles explosions sont bien plus fréquentes qu'on l'avait cru, et que certaines se produisent régulièrement dans le centre de notre galaxie. Il a montré aussi que les explosions du cœur des galaxies produisent des ondes électromagnétiques et des faisceaux de particules cosmiques particulièrement intenses qui peuvent se propager jusqu'à des distances lointaines sans être arrêtées.
Les super-ondes issus du cœur de notre galaxie peuvent donc facilement affecter la Terre. L'examen des annales contenues dans les carottes de glace polaires de la Terre (teneur en béryllium 10) indiquent que les super-ondes majeures se produisent tous les 26'000 ans (à 3000 ans près), mais qu'elles peuvent aussi se succéder après seulement 10'000 à 13'000 ans.
Des données astronomiques et géologiques révèlent qu'une super-onde issues du centre galactique a impacté notre système solaire il y a entre 16'000 et 11'000 ans, soit à la fin de la dernière ère glaciaire. Pendant ce temps, l'activité des tache solaires a été hautement renforcée.
L'influence de la super-onde est le scénario le plus probable pour expliquer plusieurs événements géophysiques survenus sur la Terre, par exemple l'extinction massive des grands mammifères de l'Amérique du Nord il y a 12'500 ans et la brusque fin de l'ère glaciaire.
Certains chercheurs pensent qu'il existe une 9e planète dans le système solaire. Encore indétectée, elle est nommée planète X. Les astronomes Konstantin Batygin et Mike Brown de l'Institut de technologie de Californie (Caltec) ont détecté des anomalies orbitales dans des astres de la ceinture de Kuiper, qu'ils attribuent à cette planète X.
Si cela est vrai, ils ont calculé qu'elle aurait une masse 10 fois plus grande que celle de la Terre, et son orbite serait 1000 fois plus éloignée du soleil, avec une période de révolution de 10 à 20'000 ans.
Les astrophysiciens états-uniens Daniel Wiltemire et John Matese ont émis l'idée que l'orbite de la planète X s'incline lentement et passe tous les 27 Ma dans la ceinture de Kuiper, une concentration de météorites s'étendant au-delà de Neptune. Cela provoquerait une expulsion de comètes dans le système solaire.
Une autre hypothèse, proposée par l'états-unien Richard A. Muller, est l'existence d'une étoile compagne du soleil, une naine brune ou une naine rouge, à l'image de nombreux systèmes d'étoiles doubles. Cette compagne est souvent nommée Némésis. (Measurement of the lunar impact record for the past 3.5 b.y. and implications for the Nemesis theory, Richard A. Muller, Geological Society of America Special Paper, 2002, 356, 659)
Némésis aurait une orbite autour du soleil située à 3 années-lumière, avec un temps de révolution de 26 Ma. Lorsqu'elle passe près du nuage d'Oort, elle déclancherait une pluie de comètes.
Le téléscope spatial WISE de la NASA, chargé d'une exploration dans le domaine infrarouge, mis en orbite en 2010, a découvert près de 35'000 astéroïdes. À ce jour, il n'a pas confirmé l'existence de Némésis ni de la planète X.
Sources:
Did a gamma-ray burst initiate the late Ordovician mass extinction? A. Melott et al., 2004, Int. J. Astrobiol. 3, 55
Gamma-ray
bursts as a threat to life on Earth, B.C. Thomas, 2009, Int. J. Astrobiol. 8, Special Issue 3, 183
Possible Role of Gamma Ray Bursts on Life Extinction in the Universe, Tsvi Piran and Raul Jimenez, Phys. Rev. Lett. 2014, 113, 231102
An ∼60 Million Year Periodicity Is Common to Marine 87Sr/86Sr, Fossil Biodiversity, and Large-Scale Sedimentation: What Does the Periodicity Reflect? Adrian L. Melott, Richard K. Bambach, Kenni D. Petersen, John M. McArthur, The Journal of Geology, 2012, 120, 2, 217;
Analysis of periodicity of extinction using the 2012 geological time scale, Adrian Melott, Richard Bambach, Paleobiology, 2014, 40, 177
Periodic variation of Oort Cloud flux and cometary impacts on the Earth and Jupiter, P. Nurmi, M.J. Valtonen and J.Q. Zheng, Mon. Not. R. Astron. Soc. 2001, 327, 1367
Did dark matter kill the dinosaurs? The Solar System's periodic passage through a 'dark disk' on the galactic plane could trigger comet bombardments that would cause mass extinctions, Elizabeth Gibney, Nature, 7 mars 2014
Researcher Links Mass Extinctions to Planet X, University of Arkansas News, 30 march 2016
La cause des extinctions massives enfin révélée? Sputnik, 31/03/2016
Nemesis, par Richard Muller
NASA's WISE Survey Finds Thousands of New Stars, But No 'Planet X', NASA news, 7 mars 2014
Paul LaViolette, article dans The Starburst Fondation: Galactic
Cosmic Ray Volleys: A Coming Global Disaster; vidéo sous-titrée, Project Camelot
*****
Quelle aventure fantastique que ce déploiement de vie sur la Terre! La Nature fait preuve d'une grande inventivité de création!
Mais nous n'en connaissons probablement qu'un tout petit aspect. D'autres découvertes nous attendent, n'en doutons pas. Elles pourraient changer notre histoire. Les mystères se dévoilent peu à peu.
Un autre aspect fascinant de la Nature dont il faudra tenir compte est que toutes les créations de l'Univers, de l'atome aux groupes de galaxies, sont sous-tendues par des trames géométriques, y compris les plantes, les animaux et les humains. (voir mon article: Aspect géométrique de l'Univers)
Reste à comprendre pourquoi la vie a évolué de cette façon. Y a-t-il une finalité, un but, un sens?
Dans le chapitre suivant (L'évolution de la Vie et les champs d'information), j'examinerai cette question, en étudiant la proposition du darwinisme et d'autres propositions bien plus intéressantes. Je répondrai que la proposition qui me semble la plus cohérente et la plus pertinente est celle qui énonce:
OUI, il y a un sens à la vie, nous y contribuons.
Et c'est magnifique!
(voir aussi Le sens de la Vie)
Autres articles de la série Une autre histoire de la Terre
31. Darwin et la théorie moderne de l'évolution. Les archives archéologiques montrent que les espèces animales et végétales ont évolué au cours des siècles. Quel est le moteur de cette évolution? Darwin et les néodarwinistes énoncent que des variations ont lieu par hasard et que certaines sont avantagées et se développent au détriment des autres. C'est ce que nous sommes conditionnés à croire (enseignement scolaire, musées, médias, etc.). Froide et aveugle, fondée sur l'égoïsme et le conflit permanent de tous contre tous, cette théorie enlève tous sens à la vie, et a donné lieu à des perversions sociales. Elle oublie que l'altruisme et l'unité de toute chose sont des principes fondamentaux de la vie.
32. Le darwinisme en question. De plus en plus de chercheurs soulignent les inconsistances de la théorie néodarwiniste, construite sur une interprétation controversée des découvertes archéologiques. L'un des points les plus contestables est que des mutations aléatoires puissent conférer des caractères avantageux. Un autre est que les caractères complexes devraient apparaitre progressivement. Or les bactéries incluent des systèmes biologiques très élaborés qui sont apparus tout faits. Les nombreuses avancées de la biologie montrent que la transmission génétique est bien plus complexe que par la voie unique des gènes. Les zones dites non-codantes jouent un rôle très important dans la régulation génétique et des instructions nouvelles peuvent passer dans la descendance sans qu'il y ait de mutations dans les gènes. Enfin, la composition de l'ADN des mitochondries montre que les espèces sont génétiquement bien distinctes, sans chainon intermédiaire. La synthèse évolutive étendue tient compte de cette complexité et de cette mobilité en intégrant l'épigénétique et la théorie des systèmes.
33. Pratiques inquiétantes dans la science. Certains scientifiques et philosophes osent exprimer leur conviction de l'existence d'un principe organisateur supérieur qui guiderait les créations des organismes vivants. Le prix à payer est de faire la séparation dans leur esprit entre un domaine subjectif qui relève de leur foi et un domaine scientifique occupé par la théorie de l'évolution. Les chercheurs qui avancent des modèles scientifiques différents sont censurés, ridiculisés, ou évincés des institutions, bien loin d'un dialogue honnête qu'on pourrait attendre de la part de scientifiques éduqués. Alors qu'elle affirme abusivement être un fait objectif, la théorie synthétique de l'évolution est en réalité une hypothèse fondée sur des présupposés métaphysiques.
34. Informations et mythes de création. La constitution des organismes, en particulier de la molécule d'ADN avec ses codes génétiques précis et volumineux, n'a pas pu avoir lieu sans l'apport extérieur d'instructions spécifiques. Elles sont fournies sous la forme de champs d'information ou archétypes, de nature électromagnétique, gravitationnelle, ou d'une nature plus subtile (éther, champs de torsion, entéléchie, champs morphogénétiques, champs H, champ akashique). Par ces champs, les êtres vivants sont reliés subtilement dans un vaste réseau. Depuis toujours, les mythes de création planétaires décrivent un processus semblable. D'un principe organisateur, émanent des instructions sous forme de pensées, de rêves, de géométrie ou de sons. Cette vision renouvelée donne à l'évolution des formes de vie un sens bien plus exaltant et plus responsable: accompagner l'éveil de la conscience.
Annexe
Classer les organismes, c'est d'abord les comparer, puis les associer par ressemblance.
Après avoir décrit de nombreux organismes animaux et végétaux, il est naturel pour un naturaliste de vouloir faire la liste de leurs ressemblances et de leurs différences, et de les rassembler par groupes.
Mais quelles sont les ressemblances qui vont être choisies? Il y a plusieurs possibilités. Devant un amoncellement de livres, un bibliothécaire doit les trier pour les ranger. Or il peut les trier par ordre alphabétique d'auteur, ou par thèmes, ou encore par année de publication. Qu'est-ce qui va guider son choix? C'est la commodité en fonction de l'usage qui en sera fait. Ce sera donc fonction de critères locaux.
En ce qui concerne les êtres vivants, la classification des plantes avant Carl von Linné (naturaliste suédois, 1707 - 1778) était utilitaire. C'était le même type que sur les catalogues de vente actuel: plantes d'ornement, arbres, arbustes, plantes alimentaires, etc..
Ensuite, de nombreuses classifications ont été proposées sur la base de l'anatomie comparée. Mais de l'organe à la molécule, en passant par les systèmes anatomiques, physiologiques, et le mode de vie, quels éléments de comparaison choisir en priorité? Cela relève toujours d'un choix subjectif arbitraire. C'est pourquoi les systèmes de classification varient selon les auteurs. Ces choix dépendent de leur sensibilité, de leur mentalité, de leur vécu.
C'est aussi pourquoi les classifications sont sujettes à des révisions fréquentes. Ainsi les tarsiers présentent des traits communs avec les anthropoides, et on pourrait donc les regrouper. Mais ils présentent également d'autres traits communs avec les omomyidés. Alors avec qui les regrouper?
Pour classer les Primates, on a retenu quelques caractéristiques communes. Mais leur classification varie selon les auteurs (voir Hominoïdes, dans le site Hominidés).
Pour définir la famille des Équidés, on a retenu les caractères des dents à couronne haute et croissance continue (hypsodontes), les membres longs, et les pieds à un seul doigt. Malgré cela, on y a inclus des animaux anciens considérés comme leurs ancêtres, alors qu'ils avaient 2 ou 3 doigts. Alors, au final, quelle est la définition des Équidés?
La subjectivité apparait même au niveau de la définition d'un règne. Il y a quelques dizaines d'années, on apprenait qu'il existe 3 règnes: minéral, végétal et animal. Car il est facile de distinguer une plante fixée au sol d'un animal mobile, et nous classons spontanément les champignons dans les plantes.
Mais les scientifiques veulent des critères précis. Mais on trouve des animaux qui sont fixés au sol, et des plantes qui sont mobiles. Bien plus, on a défini les végétaux comme ayant la capacité de photosynthèse. Celle-ci est assurée par la présence d'organites nommées chloroplastes dans les cellules (voir schéma de la cellule en début d'article). Or les champignons n'en ont pas. Ils ont donc été classés comme un règne à part en 1969. D'autres considérations ont fait augmenter le nombre de règnes à 7 actuellement
Avec les théories de l'évolution, la méthode de classification retenue actuellement tient compte de la parenté avec des espèces du passé. La dimension temporelle y prend la place centrale. C'est le classement phylogénétique ou cladistique, proposé dès 1965 par le biologiste allemand Willi Hennig (1913 - 1976).
Hennig part du postulat que toutes les espèces existant ensemble à une époque donnée sont issues du fractionnement de communautés plus anciennes. Un groupe monophylétique rassemble les espèces dont les membres sont considérés comme étroitement apparentés entre eux. (Phylogenetic systematics, 1965, Ann. Rev. Entom, 10, 97)
Sources: Classification scientifique des espèces dans wikipedia; La classification des animaux dans L'école du Dirlo; Le règne animal, Éducation à l'Environnement; Cranial remains of an Eocene tarsier, James B. Rossie, Xijun Ni, and K. Christopher Beard, PNAS 2006, 103, 12, 4381; Classification phylogénétique dans Wikipédia
| |
Texte conforme à la nouvelle orthographe française (1990)
16 mai 2019 - ajout 18 aout 2019 (super-onde galactique)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}